在谎言横行肆虐、不辨是非真伪的时代,摆证据、讲事实是没有市场的。
新冠病毒的来源真相,新冠疫情的发生真相不应是小圈子里的秘密。
接续:谁设计、制造了新冠病毒(四)欲盖弥彰 2
https://blog.creaders.net/u/13546/202305/463178.html
回归第四组证据本身。
新冠S蛋白与人类ENaC-α的结构巧合
早前的
谁设计、制造了新冠病毒(四)第四组证据概要
https://blog.creaders.net/u/13546/202305/461353.html
一文指出:
新冠病毒S蛋白与人类ENaC-α具有相同的长FCS序列,新冠病毒S蛋白S1亚基、S2亚基交界处(S1/S2处)的长furin切割序列,人类ENaC-α的长furin切割序列同为RRAR'SVAS。
FCS:Furin Cleavage Site,furin(弗林)切割位点;
ENaC-α:上皮钠离子通道的α亚基;
RRARSVAS:一个八氨基酸序列,序列中氨基酸的中文名依次为,精氨酸-精氨酸-丙氨酸-精氨酸-丝氨酸-缬氨酸-丙氨酸-丝氨酸。
RRAR'SVAS中间所加符号“'”表示furin蛋白酶的切割位置,在新冠病毒中,它也代表S蛋白(Spike蛋白,刺突蛋白)S1亚基、S2亚基的分界线。“'”符号前的四个氨基酸RRAR是新冠病毒的基本furin切割序列(或称最小furin切割序列),它们是S1亚基的最后四个氨基酸(SVAS是S2亚基的起始四个氨基酸)。
在新冠病毒中,RRAR‘SVAS是S蛋白aa(amino acid,氨基酸)序列的第683-690氨基酸(残基),在人类ENaC-α中,它是aa序列的第201-208氨基酸(残基),如下图所示:
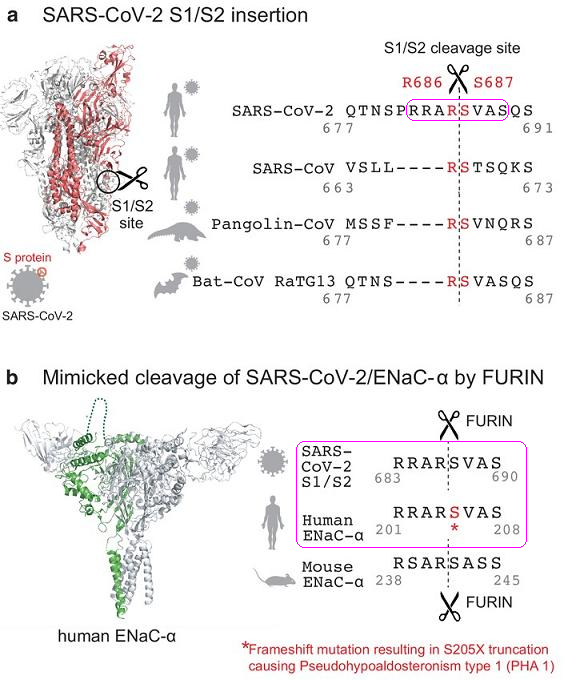
图a:新冠、三种新冠“近亲”S1/S2附近的aa序列对照。
图b:新冠S蛋白、人类ENaC-α、小鼠ENaC-α的长furin切割序列对照。
(一个与文章主旨无关的次要说明:小鼠-Mouse ENaC-α的长furin切割序列RSAR‘SASS可能应为Mouse ENaC-α的第228~235氨基酸,而不是上图所示的第238~245氨基酸。)
ENaC是Epithelial Na+ Channel(上皮钠离子通道)的缩写,也叫做Epithelial Sodium Channel (上皮钠通道),是控制钠离子通过细胞膜的跨膜离子通道。ENaC分布于人类及脊椎动物的肺(肺泡)、呼吸道(鼻腔、气管、支气管)、口腔(舌角质形成细胞)、肾脏、结肠、胰腺、汗腺、唾液腺等众多器官、结构的上皮组织中。一些ENaC高丰度分布的部位,如鼻腔上皮、上下呼吸道上皮、肺泡上皮,恰恰也是新冠病毒的首要感染部位;另有研究(论文依据一)发现,ENaC与ACE2受体在人体内的分布部位高度重合。
ENaC是由三个拓扑结构相似的亚基(亚结构)α(或δ)、β 、γ构成的三聚体。ENaC-α表示ENaC的α亚基,它有时也表示为α-ENaC或ENaC-alpha。

跨膜离子通道ENaC简单示意图
ENaC负责调节机体钠吸收,调节上皮组织表面液量,调节器官、组织的盐水平衡,调节血压等基本生理机能。ENaC变异或机能紊乱、失常可能引发各种症状,如:高血压,低血钾,肾脏水、钠潴留、代谢性碱中毒、气道表面液体体积消耗(耗竭)、粘液阻塞、纤毛清除力差、杯状细胞化生、中性粒细胞炎症等等;Liddle综合症、I型假性醛固酮减少症、(肺、呼吸道、肠道、胰脏、肝脏、肾脏、唾液腺、生殖道上皮组织的)囊性纤维化都是ENaC变异或机能异常引发的疾病。
与人类ENaC-α的长furin切割序列“巧合”使新冠S蛋白得以在细胞表面冒充、利用、竞争、劫持人类ENaC-α的水解、激活机能(ENaC的α和γ亚基需要在furin等蛋白酶作用下各自被水解切割其以激活ENaC的通道功能;完成受体结合的新冠病毒S蛋白也需要被furin等蛋白酶水解切割,S1亚基被裂解、脱落后,S2亚基将暴露在细胞膜表面,通过其融合肽介导病毒包膜与细胞膜发生膜融合,进而向细胞内释放病毒RNA),使人体细胞、人体蛋白酶将病毒结构视为人体结构,有效提升了新冠病毒的感染效率和致病能力;与人体ENaC竞争furin蛋白酶,或劫持ENaC所需的furin蛋白酶,将造成ENaC功能紊乱、失常,这一效果可能与新冠病毒的其它致病因素相互作用、相互叠加,引发更严重或更复杂的症状,譬如急性呼吸窘迫综合征(ARDS)。
由于二者切割序列完全一致,因此,对新冠S蛋白furin切割位点的治疗、阻断将也将破坏ENaC的正常功能。
(注:新冠病毒可能还存在其它模拟、模仿ENaC的结构或机制,如S蛋白中聚糖相关的结构或机制)。
新冠S蛋白与人类ENaC-α共有的RRAR'SVAS序列在(广义)生物界极为罕见,从自然演化的角度看,这一结构巧合是不可思议的,这至少体现在:
1、只有一种冠状病毒的S蛋白含有RRAR'SVAS序列,它就是新冠病毒,即,RRAR'SVAS序列在除新冠外的所有冠状病毒S蛋白中皆不存在(基于论文依据一的冠状病毒S蛋白搜索匹配结果);
注:RRAR'SVAS序列有可能在其它冠状病毒或非冠状病毒的结构蛋白中都不存在,没有人报告,在何种其它病毒的何种蛋白中也搜索匹配到了这一八氨基酸序列。
2、就人类而言,RRAR'SVAS序列只存在于ENaC-α之中,更具体点说,ENaC-α是aa(amino acid,氨基酸)序列含有RRAR'SVAS的唯一一种人体蛋白结构(基于论文依据一的人类蛋白搜索匹配结果);
3、ENaC-α含有RRAR'SVAS序列的生物物种仅有六种,其中五种是灵长类动物:人类(human)、黑猩猩(chimpanzee)、倭黑猩猩(bonobo)、猩猩(orangutan)、大猩猩(gorilla),唯一一种ENaC-α含有RRAR'SVAS序列的非人类、非类人猿物种是一种发现于欧洲和西亚的叫作pipistrellus kuhlii的蝙蝠(基于论文依据二的报告)。
4、在新冠病毒近3万个核苷酸(碱基)位点中,RRAR'SVAS序列恰巧出现在对感染、致病能力至关重要的S蛋白S1亚基、S2亚基交界处。
论文依据一:
SARS-CoV-2 strategically mimics proteolytic activation of human ENaC
(SARS-CoV-2策略性地模拟人ENaC的蛋白水解激活)
https://elifesciences.org/articles/58603
论文依据二:
A call for an independent inquiry into the origin of the SARS-CoV-2 virus
(呼吁对SARS-COV-2病毒的起源进行独立调查)
https://www.pnas.org/doi/10.1073/pnas.2202769119
新冠病毒S蛋白与人类ENaC-α共有的,在冠状病毒S蛋白中独一无二的,在生物界极为罕见的,恰巧位于新冠病毒S1、S2亚基交界处的长furin切割序列RRAR'SVAS,是自然演化的产物,还是人为赋予的?
北卡教堂山分校肺研究所
北卡罗来纳大学(UNC)教堂山分校(University of North Carolina at Chapel Hill)有一批长期研究ENaC的分子生物学-生物化学领域顶尖专家,他们所属单位是UNC-at-CH肺研究所/肺研究中心。
北卡罗来纳大学教堂山分校肺研究所/肺研究中心在不同时期名称有所不同,如在2001~2002年的一篇ENaC论文(论文-10)中,其名称为:
Cystic Fibrosis Research and Treatment Center, Department of Medicine, University of North Carolina at Chapel Hill
囊性纤维化研究与治疗中心,医学系,北卡罗来纳大学教堂山分校。
在2008年的一篇ENaC论文(论文-7)中,其名称为:
Cystic Fibrosis/Pulmonary Research and Treatment Center,University of North Carolina at Chapel Hill
囊性纤维化/肺研究和治疗中心,北卡罗来纳大纳教堂山分校。
在2018的一篇ENaC论文(论文-1)中,其名称为:
Marsico Lung Institute, Cystic Fibrosis Center and Department of Medicine, University of North Carolina, Chapel Hill, NC
Marsico肺研究所,囊性纤维化中心和医学系,北卡罗来纳大学教堂山分校。
或许您已经注意到了,北卡肺研究所三个不同时期的全名都包含“囊性纤维化”(CF,Cystic Fibrosis)一词。“囊性纤维化”是一种ENaC变异或机能异常引发的上皮组织疾病,肺泡上皮、呼吸道上皮是CF最常见的发病部位。ENaC是北卡肺研究所的核心研究领域(之一)。
北卡肺研究所对ENaC的研究
北卡教堂山分校肺研究所/肺研究中心在2001年至2018年间发表了相当数量的专门研究ENaC的论文及与ENaC高度相关的论文,下面是其中的12篇论文。
论文-1、
2018 Aug 6,Epub 2018 Jul 6(电子版在线发布时间为2018年7月6日,印刷版发行时间为2018 年8月6日)
The N terminus of α-ENaC mediates ENaC cleavage and activation by furin
α-ENaC 的N末端(氨基端,NH2端)介导弗林蛋白酶对ENaC的切割和激活
https://rupress.org/jgp/article/150/8/1179/43745/The-N-terminus-of-ENaC-mediates-ENaC-cleavage-and
https://pubmed.ncbi.nlm.nih.gov/29980634/
论文-2、
2014 Aug 15,Epub 2014 Jun 28
The N-terminal domain allosterically regulates cleavage and activation of the epithelial sodium channel
N端结构域变构调节上皮钠通道的切割和激活
https://www.jbc.org/article/S0021-9258(20)33101-X/fulltext
https://pubmed.ncbi.nlm.nih.gov/24973914/
该论文由北卡肺研究所与University of Wyoming、Medical College of Wisconsin的有关单位联合发表。
论文-3、
2012 Apr 24,Epub 2012 Apr 9
Energetic and structural basis for activation of the epithelial sodium channel by matriptase
matriptase(基质蛋白酶)激活上皮钠通道的能量和结构基础
https://pubs.acs.org/doi/10.1021/bi2014773
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3404201/
https://pubmed.ncbi.nlm.nih.gov/22471557/
该论文的发表单位除北卡肺研究所外,还包括Hotchkiss Brain Institute, Calgary, AB, Canada.
论文-4、
2012 Jan 15,Epub 2011 Oct 14
Transgenic hCFTR expression fails to correct β-ENaC mouse lung disease
hCFTR转基因表达未能纠正(改善)β-ENaC(结构、机能异常引发的)小鼠肺部疾病
https://journals.physiology.org/doi/full/10.1152/ajplung.00083.2011
https://pubmed.ncbi.nlm.nih.gov/22003093/
CFTR:cystic fibrosis transmembrane conductance regulator,囊性纤维化跨膜传导调节蛋白(调节器、调节剂)。
hCFTR:human CFTR,人类囊性纤维化跨膜传导调节蛋白。
Transgenic hCFTR expression:以hCFTR替换Mouse的相应蛋白。
论文-5、
2010 Oct 15,Epub 2010 Aug 13
The Cystic Fibrosis Transmembrane Conductance Regulator Impedes Proteolytic Stimulation of the Epithelial Na+ Channel
囊性纤维化跨膜电导调节剂阻碍上皮Na+通道的蛋白水解刺激
https://www.jbc.org/article/S0021-9258(20)47271-0/fulltext
https://pubmed.ncbi.nlm.nih.gov/20709758/
论文-6、
2010 Aug 27,Epub 2010 Jun 21
Airway surface liquid volume regulation determines different airway phenotypes in liddle compared with betaENaC-overexpressing mice
相比β-ENaC过表达小鼠,对liddle(综合症小鼠或患者)而言,不同的呼吸道表型由呼吸道表面液体容量调节决定。
https://www.jbc.org/article/S0021-9258(20)59487-8/fulltext
https://pubmed.ncbi.nlm.nih.gov/20566636/
论文-7、
2008 Nov 1,Epub 2008 Oct 13
ENaC proteolytic regulation by channel-activating protease 2
通道激活蛋白酶2(CAP2/TMPRSS4)对ENaC的蛋白水解调节
https://rupress.org/jgp/article-lookup/doi/10.1085/jgp.200810030
https://pubmed.ncbi.nlm.nih.gov/18852303/
论文-8、
Published online 1 May 2005,Published in print 1 May 2005,First published January 7, 2005
Neutrophil elastase activates near-silent epithelial Na+ channels and increases airway epithelial Na+ transport
中性粒细胞弹性蛋白酶激活近静默上皮Na+通道并增加气道上皮Na+转运
https://journals.physiology.org/doi/full/10.1152/ajplung.00435.2004
https://pubmed.ncbi.nlm.nih.gov/15640288/
论文-9、
Published online 1 January 2004,Published in print 1 January 2004,First published September 10, 2003
Serine protease activation of near-silent epithelial Na+ channels
丝氨酸蛋白酶激活近静默上皮Na+通道
https://journals.physiology.org/doi/full/10.1152/ajpcell.00342.2003
https://pubmed.ncbi.nlm.nih.gov/12967915/
论文-10、
2002 Mar 8,Epub 2001 Dec 26
Regulation of the epithelial sodium channel by serine proteases in human airways
丝氨酸蛋白酶对人气道中上皮钠通道的调节
https://www.jbc.org/article/S0021-9258(19)36437-3/fulltext
https://pubmed.ncbi.nlm.nih.gov/11756432/
以上论文都是专门研究ENaC微观结构与内在机能的分子生物学/生物化学论文;除上述10篇论文外,北卡肺研究所还发表过一些与ENaC高度相关的论文,下面是两个例子(笔者未搜集更多这类论文):
论文-11、
2006 May,Epub April 24 2006
Soluble Mediators, Not Cilia, Determine Airway Surface Liquid Volume in Normal and Cystic Fibrosis Superficial Airway Epithelia
可溶性介质,而不是纤毛,决定正常和囊性纤维化浅表气道上皮的表面液体体积
https://rupress.org/jgp/article/127/5/591/42642/Soluble-Mediators-Not-Cilia-Determine-Airway
https://pubmed.ncbi.nlm.nih.gov/16636206/
论文-12、
2010 Mar,Epub 2009 Dec 11
Modulation of endocytic trafficking and apical stability of CFTR in primary human airway epithelial cultures
原代人气道上皮培养物中CFTR内吞运输和顶端稳定性的调节
https://journals.physiology.org/doi/full/10.1152/ajplung.00016.2009
https://pubmed.ncbi.nlm.nih.gov/20008117/
分子生物学、生物化学界对RRAR'SVAS的认知
2004年Pittsburgh、Texas大学论文
2004 Apr 30(Epub 2004 Mar 7),University of Pittsburgh School of Medicine(匹兹堡大学医学院)、University of Texas Health Science Center(德克萨斯大学健康科学中心)联合发表了如下JBC(Journal of Biological Chemistry)论文:
Epithelial Sodium Channels Are Activated by Furin-dependent Proteolysis
上皮钠通道被弗林蛋白酶依赖性蛋白水解激活
https://www.jbc.org/article/S0021-9258(19)75354-X/fulltext
https://pubmed.ncbi.nlm.nih.gov/15007080/
这篇论文指出了furin蛋白酶对ENaC水解激活的重要性。
论文“Supplementary Material”部分提供了如下文件:
https://www.jbc.org/cms/10.1074/jbc.C400080200/attachment/4e4a51a9-8c96-4c20-aa59-7a96df0ee6de/mmc1.pdf
文件标出了几个物种ENaC 的α和γ亚基的所有基本/最小(minmal)furin切割序列。

几个物种ENaC的furin切割序列(ENaC-α部分)
上图是文件截图的上半部分(alpha亚基部分)。由图可见,human ENaC alpha亚基第199-204氨基酸是GARRAR,即第201-204氨基酸为RRAR(图中所有基本furin切割序列RXXR的首尾两个精氨酸皆加黑突出显示)。RRAR也正是新冠病毒S1/S2位点处的基本furin切割序列。
这表明,不晚于2004年,分子生物学、生物化学界就已经知道human ENaC-α的(最后一个)基本furin切割序列是RRAR(另二个基本furin切割序列是RSRR、RDLR);文件虽未完整展示human ENaC-α的长furin切割序列RRAR'SVAS,但分子生物学家、生物化学家们不会不知道紧随RRAR之后的四个氨基酸是SVAS。
这篇Pittsburgh、Texas大学2004年论文是论文-7(2008 Nov 1 北卡ENaC论文)的参考论文之一,论文-7在涉及furin切割的相关内容中,四次提及这篇论文,并引用论文的有关结论、发现。这篇2004年论文也被论文-1(2018 Aug 6 北卡ENaC论文)、论文-5(2010 Oct 15 北卡ENaC论文)、论文-8(1 May 2005北卡ENaC论文)列为参考论文。
2007~2008年Pittsburgh、Tennessee大学论文
01 Jan 2008(Epub 2007 Nov 21),University of Pittsburgh、University of Tennessee 联合发表了如下论文:
Defining an inhibitory domain in the α-subunit of the epithelial sodium channel
在上皮钠通道的α亚基中定义抑制域
https://journals.physiology.org/doi/full/10.1152/ajprenal.00399.2007
https://pubmed.ncbi.nlm.nih.gov/18032549/
下面是论文的Fig. 5(笔者加了若干标记方框)

Fig. 5 部分哺乳动物α-ENaC的furin切割序列
在Fig. 5中,α-ENaC包含“RRAR”切割序列的物种有四种:Oryctolagus cuniculus(穴兔或家兔)、Homo sapiens(智人或人类)、Canis familiaris(犬类)、Cavia porcellus(豚鼠)。
(注:按图中所标的氨基酸序号,Homo sapiens的α-ENaC的RRAR序列为第202-205氨基酸,而在上一个图中,human ENaC alpha的RRAR序列为第201-204氨基酸,二者有一个序号错位。)
Fig. 5也未展示RRAR之后的氨基酸。
哥伦比亚大学杰弗里·萨克斯教授(Jeffrey D. Sachs)、尼尔·哈里森(Neil L. Harrison)教授2022年5月19日发表的PNAS(Proceedings of National Academy of Sciences,美国国家科学院院刊)文章(即论文依据二):
A call for an independent inquiry into the origin of the SARS-CoV-2 virus
(呼吁对SARS-COV-2病毒的起源进行独立调查)
https://www.pnas.org/doi/10.1073/pnas.2202769119
指出:
The FCS of human ENaC α has the amino acid sequence RRAR'SVAS (Fig. 2), an eight–amino-acid sequence that is perfectly identical with the FCS of SARS-CoV-2 (16).
...
The FCS sequence of human ENaC α (20) is identical in chimpanzee, bonobo, orangutan, and gorilla (SI Appendix, Fig. 1), but diverges in all other species, even primates, except one. (The one non-human non-great ape species with the same sequence is Pipistrellus kuhlii, a bat species found in Europe and Western Asia;
...
人类ENaC α的FCS(furin cleavage site,furin切割位点)具有氨基酸序列RRAR'SVAS,这一八氨基酸序列与SARS-CoV-2的FCS(序列)完全相同。
。。。
人类ENaC α 的FCS序列(指RRAR'SVAS)与黑猩猩、矮黑猩猩、猩猩和大猩猩相同,但与所有其它物种,甚至其它灵长类动物都不相同,除了一个例外。(这一非人类、非类人猿类的具有相同序列的唯一例外物种是Pipistrellus kuhlii,一种发现于欧洲和西亚的蝙蝠;
。。。
由以上内容可知,ENaC-α含有RRAR'SVAS序列的物种仅有六种:人类(human)、黑猩猩(chimpanzee)、倭黑猩猩(bonobo)、猩猩(orangutan)、大猩猩(gorilla)、Pipistrellus kuhlii蝙蝠。
进而可知,在Fig. 5中,除Homo sapiens(智人或人类)外,其它三个ENaC-α含有基本furin切割序列RRAR的物种(Oryctolagus cuniculus、Canis familiaris、Cavia porcellus),其RRAR之后的四个氨基酸不会是SVAS。
2007~2008年的Pittsburgh大学、Tennessee大学论文再次表明,人类ENaC-α的furin切割序列是分子生物学界、生物化学界的一个“常识”,而且,分子生物学家、生物化学家们经常比对human与其它物种ENaC的furin切割序列。
2016年Israel Ariel、Tel-Aviv大学论文
2016 Apr 1(Epub 2016 Jan 7), Israel Ariel University、Tel-Aviv University的两位科学家发表了如下论文:
Epithelial sodium channel (ENaC) family Phylogeny, structure-function, tissue distribution, and associated inherited diseases
上皮钠通道(ENaC)家族系统发育、结构-功能、组织分布,以及相关遗传疾病
https://www.sciencedirect.com/science/article/abs/pii/S0378111915015735
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4756657/
https://pubmed.ncbi.nlm.nih.gov/26772908/
论文Fig. 11的α亚基部分如下:

Fig. 11 20个物种胞外域的丝氨酸蛋白酶切割位点(ENaC-α部分)
(注:furin蛋白酶是一种前体蛋白转化酶,而前体蛋白转化酶又是丝氨酸蛋白酶大家族中的一个子类。简而言之,furin蛋白酶是一种丝氨酸蛋白酶;Fig. 11还涉及另一种丝氨酸蛋白酶,胰蛋白酶-Trypsin,它以单个的精氨酸-R为切割位点。)
在Fig. 11所列20个物种中,ENaC-α包含RRAR'SVAS序列的是前三个物种(human-人类,chimpanzee-黑猩猩,Gorilla-大猩猩)。
(注:按Fig. 11所标的氨基酸序号,RRAR'SVAS序列是human ENaC-α的第208-215氨基酸,而不是前文所说的第201-208氨基酸。笔者猜测,该论文对human ENaC-α起始氨基酸的定义可能与前面几篇相关论文不同。)
由上述三篇论文可知,分子生物学、生物化学界早就知道,非常清楚人类ENaC-α的长furin切割序列RRAR'SVAS。
新冠病毒S蛋白与人类ENaC-α共有的,在冠状病毒S蛋白中独一无二的,在生物界极为罕见的,恰巧位于S蛋白S1/S2位点处的长furin切割序列RRAR'SVAS,是自然演化的产物,还是人为设计、赋予的?
新冠病毒是专为人类设计的。
谁将人类ENaC-α的长furin切割序列RRAR'SVAS引入到了新冠病毒S蛋白S1亚基、S2亚基交界处?是分子生物学家、生物化学家?是北卡肺研究所?谁才是新冠病毒的核心设计者?
(未完待续)
| 

