接续:
谁设计、制造了新冠病毒(二 中)
谁设计、制造了新冠病毒(二 中)续
新冠病毒S1/S2处(S蛋白S1、S2亚基交界处)的“RRAR”furin酶切位点不是自然演化(自然变异或自然重组)产生的,它是借鉴MHV-JHM(一种小鼠肝炎病毒)、MHV-3、FIPV(猫传染性腹膜炎病毒)等高致病性、高致死率病毒S1/S2处的“RRARR”furin酶切位点,人为设计、人为引入的。
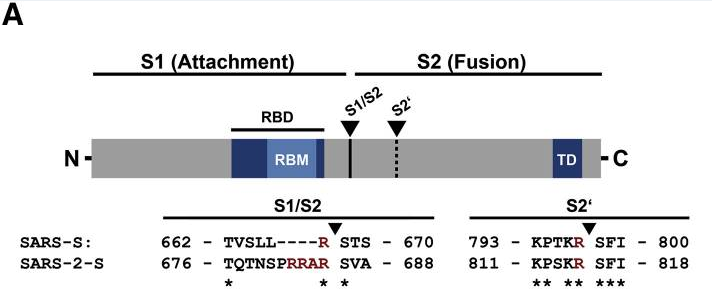
新冠病毒S1/S2处的“PRRA”插入及“RRAR”furin酶切位点
谁设计了新冠病毒的furin酶切位点?
本文将展示多组证据,这些证据指示着同一个结论:新冠病毒S1/S2处的“RRAR”furin酶切位点,极有可能是北卡罗来纳大学教堂山分校微生物学和免疫学院流行病学系教授,冠状病毒“合成之父”,冠状病毒功能增益改造狂人,现美国国家科学院院士Ralph
S. Baric(拉尔夫·S·巴里克)设计、引入的。
不只是furin酶切位点,Ralph S. Baric的研究内容、论文还与新冠病毒的其它多项结构、特性存在极为直接的重大关联。
由“新冠病毒与拉尔夫·巴里克的不解之缘(一)”一文可知,Ralph S. Baric也是新冠病毒“逆转录RNA为cDNA并整合入人体DNA”特性的第一设计嫌疑人;
由“连系新冠病毒与Ralph S. Baric的双纽带(上)、(中)、(下)”等文章可知,Ralph S. Baric同时还是新冠病毒RBD关键氨基酸(决定冠状病毒的ACE2结合能力、细胞进入能力和宿主范围)的最大设计嫌疑人。
由“新冠病毒溯源八问(上)”等文章可知,新冠病毒还有一组独一无二的能力,它集免疫破坏、免疫屏蔽、免疫逃避、免疫抑制、免疫干扰等宿主免疫对抗机制于一身,没有任何一种其它冠状病毒,没有任何一种非冠状病毒具备如此全面的免疫对抗机制!这是新冠病毒又一组极为明显的人为设计特征,Ralph
S. Baric也极有可能是这组免疫对抗机制的设计者或联合设计者,稍后我们将展示具体的证据:在生态健康联盟2018年组织的Defuse
Project中,Ralph S. Baric(团队)不仅负责furin酶切位点插入研究,而且同时还领导靶向免疫增强工作(Prof.
Baric, Univ.N.Carolina, will lead targeted immune boosting work)。
Ralph
S.
Baric与新冠病毒诸多结构、特性的重大关联(furin酶切位点、逆转录RNA为cDNA并整合入人体DNA、RBD关键氨基酸、全面的宿主免疫对抗机制等等),不可能全都是巧合,新冠病毒也不可能独独与Ralph
S. Baric一人同时发生如此之多的巧合。它们不是巧合,它们是如下结论的铁证:Ralph S. Baric就是新冠病毒的核心设计者!
下面将展示五组furin酶切位点相关的证据。与新冠病毒其它结构、特性相关的,指证Ralph S. Baric设计、制造新冠病毒的证据,已在或将在本系列其它文章中展示。
I Ralph S. Baric、小鼠肝炎病毒、“RRARR”酶切位点
不晚于2000年11月,Ralph S. Baric领导的小组发明了基于基因序列合成冠状病毒的反向遗传平台,并使用该平台合成了猪传染性胃肠炎病毒TGEV(Transmissible GastroEnteritis Virus)。
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC110934/
这是科学界首次人工合成冠状病毒。TGEV是一种α谱系冠状病毒。
反向遗传平台的基本功能特点是,只要知道冠状病毒的基因序列,就可合成序列对应的冠状病毒cDNA、RNA,并通过病毒RNA在培养细胞中复活病毒毒株。用于合成病毒的基因序列可以是已知冠状病毒的基因序列,也可以是人为设计的基因序列(甚至可以是用多个基因序列拼合出的基因序列)。反向遗传平台不仅可以克隆已知冠状病毒,还可以无痕迹地灵活改造冠状病毒,它一定程度上将冠状病毒的改造工作简化为基因序列文本的编辑、设计(替换、修改、拼装)。
2002年11月前,Rralph S. Baric 领导的一个小团队第二次使用反向遗传平台合成冠状病毒,这一次合成的是MHV-A59(小鼠肝炎病毒A59株系)。
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC136593/
MHV(Mouse Hepatitis Virus,小鼠肝炎病毒)属于冠状病毒β谱系A支系,新冠病毒(、SARS病毒及众多SARS-like蝙蝠冠状病毒)属于β谱系B支系。MHV-A59是一个毒性较强的MHV株系。
MHV-A59和新冠病毒有一个结构共同点:它们S蛋白S1/S2处都有Furin酶切位点。MHV-A59
S1/S2处的复合酶切组合是“RRAHR”(精氨酸-精氨酸-丙氨酸-组氨酸-精氨酸,包含基本furin酶切组合“RAHR”,furin酶切组合的模式是“RXXR”),新冠病毒S1/S2处的furin酶切组合是“RRAR”。
小鼠是最常用的实验室动物,小鼠肝炎病毒(MHV)是科学界研究最频繁的动物病毒。小鼠肝炎病毒有超过25个株系,其中最受关注、被研究最多的三个株系是MHV-JHM(也叫MHV-4)、MHV-3、MHV-A59。MHV-JHM和MHV-3是毒性最强、次强的MHV株系,它们对BALB/c小鼠(白变种实验室小鼠)的感染致死率接近100%。
MHV-JHM、MHV-3的强大致病能力与它们的酶切结构有关。这两种病毒S1/S2处都有一个“RRARR”(精氨酸-精氨酸-丙氨酸-精氨酸-精氨酸)酶切位点,这一复合酶切组合包含两个基本furin酶切组合“RRAR”和“RARR”。非常“巧合”的是,前者“RRAR”恰恰就是新冠病毒S1/S2处的furin酶切组合。
在β谱系B支系的众多冠状病毒中,S1/S2处有furin酶切位点的病毒只有一种,就是新冠病毒。也就是说,新冠病毒S1/S2处的furin酶切位点结构在其近亲病毒中是独一无二的。
在新冠病毒出现前,S1/S2处具有“RRAR”或“RRARR”酶切位点的冠状病毒有三种(类),除MHV-JHM、MHV-3外,某些猫传染性腹膜炎病毒(Feline
Infectious Peritonitis
Virus,FIPV)病毒株S1/S2处的复合酶切组合也是“RRARR”。这些FIPV病毒对猫是极其致命的,一年内的感染致死率几乎绝对100%。MHV与新冠病毒同处冠状病毒β谱系,但分属A、B两支系;FIPV与新冠病毒的关系更远,它属于冠状病毒α谱系,与前面提到的猪传染性胃肠炎病毒TGEV同谱系。
虽然同属冠状病毒β谱系,但MHV与新冠病毒亲缘关系极远,结构差异极大。
注:
新冠与MHV-JHM,
二者全基因组序列仅有23%具有可比对性(即二者77%的序列几无相似之处),23%的可比对部分的一致性仅为66.79%;
二者S蛋白aa(amino acid,氨基酸)序列92% 可比对,但可比对部分的一致性仅为36.65%。
新冠与MHV-3,
二者全基因组序列仅有24%可比对,24%可比对部分的一致性仅为66.27%;
二者S蛋白aa序列91%可比对,但可比对部分的一致性仅为37.42%。
亲缘关系极远,结构差异极大的MHV-JHM、MHV-3
、FIPV
S1/S2处的“RRARR”酶切组合中的“RRAR”为什么会出现在新冠病毒S1/S2处?这不是自然演化的结果,不是自然变异的巧合;有人借鉴了这三种病毒的强力致病结构,将之设计、应用到了新冠病毒中。
使用反向遗传平台合成冠状病毒是基于基因序列的。合成过MHV-A59的Ralph
S. Baric对其 S1/S2处的furin酶切位点肯定一清二楚;对毒性最强,知名度不亚于MHV-A59的MHV-JHM、MHV-3
这两种小鼠肝炎病毒S1/S2处的“RRARR”酶切位点及其强大致病作用,Ralph S. Baric肯定也了如指掌。
当然,MHV-A59、MHV-JHM、MHV-3三种病毒,及后二种病毒S1/S2处的“RRARR”酶切位点在病毒学界是广为人知的,可能借鉴“RRARR”并将其中的“RRAR”设计、应用到新冠病毒S1/S2处的病毒学家大有人在,不只Ralph
S. Baric一人。
我们需要看进一步的证据。
II Baric2016年对未来病毒酶切特性的预测
2016年3月14日,Ralph
S. Baric领导的团队在PNAS(Proceedings of the National Academy of Sciences of
the United States of America,美国国家科学院院刊)上发表了如下论文:
SARS-like WIV1-CoV poised for human emergence
(类SARS冠状病毒WIV1-CoV有产生人类流行疫情的潜在危险)
https://www.pnas.org/content/113/11/3048
奥巴马功能增益研究暂停令(2014年10月17日)颁布后,经Ralph
S.
Baric要求,其团队两项SARS相关的功能增益改造研究得到NIH(美国国立卫生研究院)特批,获准继续实施,其中一项(嵌合)改造研究后来发表为上述2016年3月的PNAS论文;另一项嵌合改造研究则发表为更为著名的2015年11月9日的Nature
Medicine论文:
https://www.nature.com/articles/nm.3985
有必要对2015年11月的Nature Medicine论文多罗嗦几句。
由于石正丽及其团队成员葛行义列名了这一论文的作者,该论文常被指鹿为马,说成是石正丽团队发表的论文。实际情况如何呢?应Ralph
S.
Baric的要求,石正丽为该研究提供了嵌合的原材料之一--SHC014(一种有ACE2结合能力和人体细胞进入能力的特殊蝙蝠冠状病毒)的S蛋白基因序列;同时,葛行义向Baric提供了2013年10月30日石正丽团队Nature论文(葛行义为论文第一作者)
https://www.nature.com/articles/nature12711
中的一项WIV1(另一种有ACE2结合能力和人体细胞进入能力的特殊蝙蝠冠状病毒)假病毒实验资料,该实验被Ralph
S.
Baric的2015年论文援引为SHC014病毒实验的对照实验)。因为提供了病毒原材料和2013年的实验资料,葛行义、石正丽分别被(礼节性地)列名为2015年论文的第9作者和第14作者(该论文最末位最次要的作者)。实际上,石正丽、葛行义只是提供了原材料和过往的实验资料,二人并未参加该项嵌合改造研究。
回归2016年3月的PNAS论文。该研究以WIV1的S蛋白和SARS-CoV MA15的骨架为原材料,基于两者基因序列的嵌合序列,使用反向遗传平台合成了一种嵌合病毒WIV1-MA15,同时合成了WIV1的病毒克隆,论文称之为WIV1-CoV。
注:2015、2016两篇论文都是用SARS-CoV MA15的骨架与蝙蝠冠状病毒的S蛋白做嵌合,2015年论文用的蝙蝠冠状病毒是SHC014,2016年论文用的是WIV1。
关于WIV1的一些背景说明。WIV1(及SHC014)是少数几种有ACE2结合能力即人体细胞进入能力的特殊蝙蝠冠状病毒;绝大多数蝙蝠冠状病毒没有ACE2结合能力,人体细胞进入能力。WIV1、SHC014等少数蝙蝠冠状病毒虽然有人体细胞进入能力,但它们不会使人体产生临床疾病症状,并会自行从人体中消失。所有已知的蝙蝠冠状病毒都没有人体致病能力。不过,某些蝙蝠冠状病毒如WIV1能使实验室小鼠产生一定疾病症状。
还有一点需要强调,WIV1也是一种与新冠病毒高度相关的病毒,它也是新冠设计者的最重要借鉴、参照对象之一。WIV1
的S蛋白(或者说S蛋白RBD)具有强大的跨物种ACE2结合能力,它赋予了WIV1强大的跨物种细胞进入能力。新冠病毒RBD的五个关键氨基酸是参照WIV1的RBD五个关键氨基酸,通过氨基酸复用和等效替换设计而成的。这一参照设计赋予了新冠病毒极其强大的跨物种感染、传播能力。
新冠病毒RBD关键氨基酸的设计情况及与WIV1 RBD关键氨基酸的关联可参考“连系新冠病毒与Ralph S. Baric的双纽带(上)、(中)、(下)”等文章。
关于SARS-CoV
MA15的背景说明。SARS病毒对人类是致命的,但它对小鼠的致病力却很弱,SARS流行毒株不会使年轻的实验小鼠产生临床疾病症状。2007年,Ralph
S. Baric
等人在年轻BALB/c小鼠(白变种实验室小鼠)呼吸道中通过15次连续传代,培养出了一种对小鼠有强大致死力的SARS病毒小鼠适应性变异体,它可使年轻的被感染BALB/c
小鼠在3-5天内100%死亡。这一对小鼠极其致命的SARS病毒实验室变异体被命名为SARS-CoV MA15(MA:Mouse-Adapted
),其培养、产生过程发表在以下论文中:
https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.0030005
言归正传。2016年论文实验证明:
a)
WIV1-MA15、WIV1-CoV都有跨物种感染能力,它们能强烈感染灵长类动物的Vero
E6细胞(非洲绿猴肾细胞),能强烈感染人类呼吸道原代上皮细胞(HAE,human airway epithelial
cell,分离自供体肺分叉上方的气道上皮) ,并在细胞内大量复制,它们在这两类细胞中的复制滴度与SARS-CoV流行病毒株相当。
b) WIV1-MA15、WIV-CoV也能感染实验小鼠,并在小鼠气道和肺中复制,不过,它们对BALB/c小鼠的致病力远弱于SARS-CoV MA15,它们不会使较年轻的实验小鼠(10周龄)致死或体重显著减轻;
c)
然而,当将普通实验小鼠换成表达人类ACE2的转基因小鼠(其肺、脑、肝、肾和胃肠道等部位的细胞跨膜受体--鼠ACE2被转基因为hACE2,即human
ACE2)时,WIV1-CoV在小鼠肺中的复制滴度提高了100倍;同时,小鼠的症状显著加重,部分10-20周龄的被感染小鼠体重减轻超过10%,并因WIV1-CoV在大脑中的强大复制而患上了致命的脑炎,到第7天时,约50%的被感染小鼠死亡。
注:这一实验结果表明,WIV1的刺突蛋白及RBD极为适合结合hACE2。事实上,WIV1不仅刺突蛋白极为适合结合hACE2,极为适合感染人体细胞,它还兼具非常强大的跨物种细胞进入、感染能力,这是Ralph
S. Baric参照WIV1的RBD关键氨基酸设计新冠病毒RBD关键氨基酸的原因所在。
d)
嵌合病毒WIV1-MA15对小鼠的致病力强于WIV1-CoV,这说明来自SARS-CoV
MA15的病毒骨架比WIV1的病毒骨架毒性(致病能力)强(WIV1有人体细胞进入、感染能力,但没有人体致病能力,它同时只能使普通的未转基因的小鼠产生较轻疾病症状。WIV1对于功能增益改造的价值在于它的刺突蛋白和RBD,在于把它的刺突蛋白和RBD嫁接、用于有人类致病力的病毒骨架)。
e) SARS单克隆抗体可有效治疗WIV1-MA15感染,但SARS疫苗对WIV1-MA15感染没有疗效,并有显著副作用。
以上是对论文内容的顺带介绍,本文要关注的重点其实是论文“Discussion”部分中的如下文字:
Whereas
the receptor binding domain had garnered the most interest, changes in
the remaining portion of S1 as well as the S2 portion of spike may also
play a critical role in facilitating CoV infection, transmission, and/or
pathogenesis (20). Differences in these regions of spike may yield
increased protease targeting, enhanced spike cleavage, and/or expanded
tropism leading to more robust infection for the epidemic SARS strains.
译文:
尽管受体结合域(Receptor
Binding
Domain,RBD)引起了人们最大的兴趣,但S1亚基的其它部分(RBD是S蛋白S1亚基的一部分),以及Spike(刺突蛋白,S蛋白)S2亚基的变化也可能在促进CoV(coronavirus,冠状病毒)感染、传播、且/或致病中发挥关键作用。这些Spike区域的差异(自然变异造成的差异或人为改造产生的差异)可能会增强蛋白酶的靶向(性),增进刺突蛋白的切割,且/或扩大的嗜性,使流行性SARS毒株感染能力更强烈(健全)。
从功能增益研究的角度看,这段话实际上提出了增强、完善SARS病毒感染能力的病毒改造方向、建议和对改造成效果的预测,即,通过改造刺突蛋白RBD外的其它部分,包括改造S1亚基RBD外部分及S2亚基,可以获得以下改造效果:
a) 使刺突蛋白更有靶向性(针对性、目的性)地与蛋白酶发生作用;
b) 增进刺突蛋白的切割(被蛋白酶水解切割);
c) 使病毒的细胞嗜性、感染向性扩大(可感染的细胞类型范围扩大),使病毒可侵害的组织、器官范围扩大。
改造效果b)、c)的含义相对明确。效果a) “与蛋白酶作用的靶向(性)”可能有两种解读:
解读之一,是指刺突蛋白与ACE2受体的结合更有针对性。ACE2:Angiotensin Converting Enzyme2,血管紧张素转换酶2,ACE2受体本身就是细胞膜上的一个跨膜蛋白酶;
解读之二,是指使刺突蛋白可被更特定的蛋白酶水解切割,如furin蛋白酶。
令人惊奇的是,2016年论文预测的上述三项病毒改造效果,在3年后出现的新冠病毒中全都应验实现了!而且,它们都是刺突蛋白RBD区域外的结构改变所赋予的。
新冠病毒的刺突蛋白整体为三关节式铰链结构,这一结构使其能够灵活地变换姿态、变换方向,或弯曲或挺直,或收缩或伸展,或倾斜(甚至倒伏或躺卧)或直立。这样的三关节式铰链结构在其它病毒中从未报道过,新冠病毒刺突蛋白的扩展性、灵活性可能是史无前例的。
https://science.sciencemag.org/content/370/6513/203
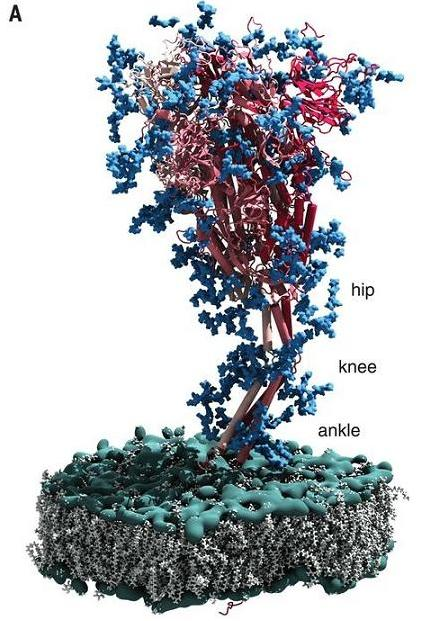
新冠病毒刺突蛋白髋、膝、踝三关节式绞链结构示意图
(三关节式铰链结构使)刺突蛋白前端的RBD平时可以处于倒伏、躺卧状态,这一姿态可以非常有效地隐蔽RBD,逃避免疫系统的搜索、攻击,逃避人类抗体或抗体药物的结合;在需要时,RBD又可以直立、暴露出来,以便接触、结合ACE2受体。这正是刺突蛋白与ACE2受体作用、结合的针对性、目的性(改造效果a的第一种解读)在新冠病毒中的应验。
https://www.pnas.org/content/117/21/11727
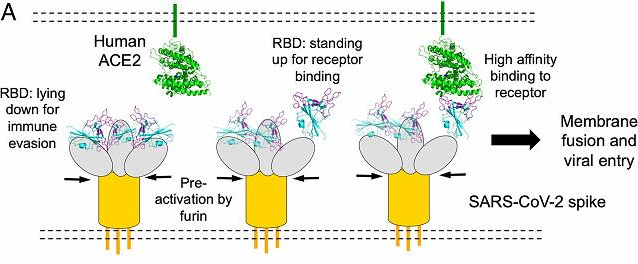
新冠病毒刺突蛋白RBD倒伏以免疫逃避(逃避抗体结合),直立以结合ACE2受体示意图
三关节式铰链结构还使新冠刺突蛋白及前端的RBD能以不同的高度、角度、姿态,灵活、多样地,更自由、高效地接触、结合ACE2受体。
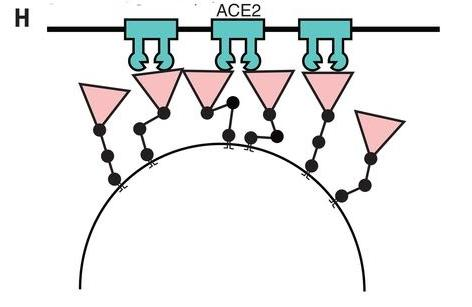
三关节式铰链使刺突蛋白(RBD)与ACE2的接合更自由、灵活、高效
由于具备S1/S2处的“RRAR”(更完整地说是“PRRAR”,“RRAR”之前的脯氨酸P可增进膜融合)furin酶切位点,完成受体结合后,新冠刺突蛋白可在细胞膜表面被furin蛋白酶水解切割;S1亚基(受体结合亚基)切割脱落后,S2亚基(膜融合亚基)得以暴露到细胞膜面前,S2亚基所含的融合肽将使病毒包膜与细胞膜发生膜融合;二膜融合后,病毒包膜内的RNA将释放到细胞内,病毒复制将第一时间开始。S1/S2处的“RRAR”(或者说“PRRAR”)酶切位点赋予新冠病毒的在细胞表面与细胞膜直接膜融合的感染方式,比之SARS病毒的内吞感染方式(先“囫囵吞枣”整体内吞到细胞中,内吞后,病毒包膜再与内吞体膜或溶酶体膜发生膜融合以释放RNA)效率高100~1000倍。
新冠病毒S1/S2处的“RRAR”或“PRRAR”furin酶切位点使2016年论文“增进刺突蛋白的切割”(效果b及效果a的解读二)的改造效果预测也在新冠中应验和实现了。
furin蛋白酶在脊椎动物的器官、组织中广泛存在,它存在的广泛性和均衡性超过了新冠病毒使用的主要受体ACE2;S1/S2处“RRAR”或“PRRAR”赋予新冠的细胞表面直接膜融合释放RNA的强悍感染效率及furin蛋白酶的广泛存在性,极大地增强、加剧了新冠病毒的泛器官、泛组织感染、扩散、侵害能力。这一结构还赋予了新冠病毒杀死淋巴细胞,破坏免疫系统的一定能力,它可能还赋予了新冠病毒破坏、穿透血脑屏障,感染、损伤大脑和神经系统的能力。这正是2016年论文改造效果c(细胞嗜性扩大)预测的应验和实现。
Ralph S. Baric2016年功能增益研究论文中的三项病毒改造预测,在新冠病毒中全都应验实现了,这难道又是巧合吗?
III Baric与Defuse项目中的furin酶切位点插入研究
2017年12月19日,川普共和党政府撤销了奥巴马禁令(奥巴马政府2014年10月17日颁布的SARS、MERS、流感相关的功能增益研究暂停令,它实际上仍然允许美国科学家通过特批开展功能增益改造研究),全面重启了美国危险病原体功能增益改造研究,重新为从事危险病原体功能增益研究的美国科学家提供联邦资金,以资助、支持相关研究。
参见:
US government lifts ban on risky pathogen research(美国政府解除了危险病原体研究禁令)
https://www.nature.com/articles/d41586-017-08837-7
大约同一时期,美国国防部高级研究计划局(Defense
Advanced Research Projects
Agency,DARPA)启动了一项名为Defuse的冠状病毒改造研究计划,将之委托给生态健康联盟(该联盟以非政府机构形式为美国政府工作,包括干脏活,是美国政府病毒研究的代理组织者,美国政府病毒研究资金的分发白手套)组织实施。2018年3月下旬,生态健康联盟将草拟完成的Defuse计划书提交DARPA审批,同时申请1400万美元的项目研究经费。
下面是生态健康联盟主席彼得·达萨克(Peter Daszak)和幕僚长阿列克谢·切姆拉(Aleksei Chmura)向DARPA提交Defuse项目计划书并申请研究经费的email截图。

Defuse项目Title全称为Project DEFUSE: defusing the threat of bat-borne coronaviruses(DEFUSE 项目:化解蝙蝠传播的冠状病毒的威胁)
必须指出,尽管少数蝙蝠冠状病毒有hACE2结合能力和人体细胞进入能力,但从未发现过有人类致病能力的蝙蝠冠状病毒。因此,并没有现实的蝙蝠冠状病毒的人类致病威胁需要化解。Defuse项目是一个冠状病毒功能增益改造研究项目,它实质上是在研究如何改造蝙蝠冠状病毒,使之具有人类致病性,使之获得各种人类致病能力;Defuse项目名为化解威胁,其实是打着预测蝙蝠冠状病毒变异的旗号主动制造人类威胁。

Defuse项目计划书截图
由上图可知:
1、Defuse是一个美国军方项目,是美国国防部启动的病毒研究计划,项目的委托、主管、出资单位是美国国防部高级计划研究局(DARPA);
2、项目的承托单位和直接组织、领导者是生态健康联盟(EcoHealth Alliance)
3、除Lead Organization生态健康联盟外,参加项目实施的Team Members还有五家,分别是:
1)、Duke-NUS Medical School
杜克-新加坡国立大学医学院,成立于2005年4月,由美国杜克大学(Duke
University)和新加坡国立大学(National University of Singapore,
NUS)联合创办。实际参加Defuse项目的是该院新发传染病研究所所长王林发(新冠自然来源论的另一个大力鼓吹者)的团队。
2)、University of North Carolina
北卡罗来纳大学。实际参加项目的是教堂山分校的冠状病毒全球头号研究权威,冠状病毒合成之父,冠状病毒功能增益改造狂人Ralph S. Baric的团队。
3)、Wuhan Institute of Virology(武汉病毒研究所)
实际参加项目的是该所的石正丽团队。
4)、USGS National Wildlife Health Center(美国地质调查局国家野生动物健康中心)
5)、Palo Alto Research Center(帕罗奥多研究中心,母公司为施乐)
以上信息表明,Defuse项目的委托、主管、出资方是美国军方;项目的承托方和直接组织、管理方是为美国政府工作的非政府组织--生态健康联盟;五个项目实施单位,三家是美国病毒研究机构,一家是美国、新加坡合办的病毒研究机构,另有一家外包研究机构--中国的武汉病毒研究所。
显而易见,Defuse是一个彻头彻尾的美国病毒研究计划,而且它还是美国军方的病毒研究计划。
Defuse多次被捂盖者、谎言者们(如在英国议会科学和技术委员会作伪证的博德研究所博士后Alina
Chan(曾昱嘉))用作栽赃、嫁祸工具,在言及Defuse的时候,他们只说这是生态健康联盟与武汉病毒研究所的合作项目,或只说武汉所是项目实施单位,对其它重要信息,他们一概隐匿不提。
新冠病毒的设计、产生与Defuse项目高度相关,新冠病毒的多项结构、特性,都与Defuse项目的研究、改造内容存在清晰的关联,本文主要聚焦其中的furin酶切位点插入研究,也将顺带提及与宿主免疫系统交互相关的研究,其它与新冠病毒结构、特性有关的项目内容可能会在后续其它文章中介绍、展开。
武汉病毒研究所在Defuse这个美国病毒研究项目中扮演着外包单位的角色。武汉所于美国病毒研究,于Defuse项目的价值在于,它是病毒改造原材料--蝙蝠冠状病毒丰富、有效提供者。我们看看武汉所在项目中的具体分工。

Defuse项目管理分工情况(计划书第1页部分内容截图)
由上图可知,武汉病毒研究所(石正丽博士团队)的项目分工是:
Dr.
Shi, Wuhan Institute of Virology will conduct viral testing on all
collected sample, binding assays and some humanized mouse work.
武汉病毒研究所石正丽博士(团队)将对采集的所有样本实施病毒检测,实施(病毒与ACE2的)结合测定,及实施一些人源化小鼠工作。
“人源化小鼠”前面提到过,就是表达人类ACE2的转基因小鼠,“人源化小鼠工作”就是用表达人类ACE2的转基因小鼠测试、鉴定蝙蝠冠状病毒的hACE2结合能力和人体细胞进入、感染能力。
作适当展开,更细致、具体点说,武汉病毒研究所在Defuse项目中的分工是:
搜集蝙蝠样本,包括蝙蝠活体样本和蝙蝠排出物样本;
检测(分离、鉴定、测序)样本中的蝙蝠冠状病毒;
测试所搜集、分离的蝙蝠冠状病毒与人或动物ACE2的结合能力(只能少数蝙蝠冠状病毒能结合蝙蝠、人类或其它物种的ACE2,通过结合ACE2受体这一途径进入、感染宿主细胞),包括用人源化小鼠测试它们与人类ACE2的结合能力,即人体细胞进入能力。
简而言之,武汉病毒研究所在Defuse项目中的分工就是搜集、分离、测序病毒原材料—蝙蝠冠状病毒,并测试它们的特性。武汉所不负责对蝙蝠冠状病毒实施功能增益改造。
再来看看北卡罗来纳大学教堂山分校Ralph S. Baric(团队)的项目分工:
Prof.
Baric, Univ.N.Carolina, will lead targeted immune boosting work, build
on his two-decade track record of reverse-engineering CoV and other
virus spike protein.
北卡罗来纳大学巴里克教授将基于他对冠状病毒及(其它?这个other 有笔误之嫌)病毒刺突蛋白二十年的反向工程经验,领导靶向(定向)免疫增强工作。
正向遗传工程是通过杂交或环境选择压力等外部手段间接改变生物基因;而反向工程(即反向遗传工程或反向基因工程)则是直接编辑、改造生物基因。常用的反向遗传(工程)基因改造手段包括基因剪刀、重叠PCR定点诱变或反向遗传平台等。
换句话说,Ralph S. Baric的分工是,通过反向遗传工程改造冠状病毒及其刺突蛋白,领导靶向免疫增强工作。
“靶向免疫增强工作”涉及病毒与宿主免疫系统的相互作用,我在计划书中尚末找到具体解释,它可能是指增强改造病毒的免疫系统对抗能力。
下图是Defuse项目计划书第12页部分内容截图,它提及了合成嵌合病毒(synthetic
chimeric viruses)、(与宿主)免疫系统交互(immune system
interaction)、(病毒在)细胞内的生长(intracellular growth)、宿主发病机制(host
pathogenesis)、宿主跳跃潜力(host jumping potential)(即跨物种感染、传播潜力)等研究内容。

Defuse最值得关注之处是,项目中有一项病毒改造内容是,在SARSr-CoV中引入furin cleavage sites (FCS,furin切割位点,即furin酶切位点)。
注:
SARSr-CoV即SARS-related-CoV,SARS相关的冠状病毒,也称SARS-like-CoV(类SAS冠状病毒),主要指位于β谱系的那些蝙蝠冠状病毒(有些蝙蝠冠状病毒位于其它谱系)。SARS病毒通常被视为冠状病毒β谱系的代表病毒,该谱系的其它病毒则被视为SARS-related-CoV或SARS-like-CoV。β谱系的主要成员是众多蝙蝠冠状病毒,同样属于β谱系的新冠病毒也是SARS-related-CoV或SARS-like-CoV。

在SARSr-Cov中引入furin cleavage sites(Defuse计划书中第11页部分内容截图)
谁负责在蝙蝠冠状病毒中引入(introduce)furin
cleavage sites?就是Ralph S. Baric。furin cleavage site
通常是刺突蛋白S1/S2处或S2亚基上的一处结构;刚刚谈过的Defuse项目分工说得很清楚,负责使用反向遗传工程改造冠状病毒及其刺突蛋白的,就是Ralph
S. Baric(团队)。
再看计划书第27页。

Defuse计划书第27页部分内容截图
注意如下内容:
Subtask
4.5 Test synthetic modifications to viral spike proteins including RBD
deletions, S2 Proteolytic Cleavage and Glycosylation Site, N-linked
glycoylation(UNC)。
子任务4.5 测试病毒刺突蛋白的合成修改,包括RBD(氨基酸)缺失、S2 蛋白水解切割和糖基化位点、N-linked糖基化 (UNC,University of North Carolina,北卡罗来纳大学) 。
可见,与多项刺突蛋白合成改造有关的测试,包括对furin酶切位点插入(即S2蛋白水解切割,插入furin酶切位点的作用就是促成furin蛋白酶对刺突蛋白进行水解切割,切割产生分离的整个S2亚基或包含融合肽的部分S2亚基)的测试,也由病毒改造团队,“furin酶切位点插入”实施团队--Ralph
S. Baric团队负责实施的。
Defuse项目中,Ralph S. Baric负责在蝙蝠冠状病毒中设计、引入furin酶切位点,而新冠病毒(新冠病毒是基于蝙蝠冠状病毒的骨架,经多重改造得到的)S1/S2处恰恰出现了“RRAR”furin酶切位点,这又是巧合吗?
Defuse项目很可能已经实施(美国方面声称DARPA没有批准Defuse项目计划书,这很可能又是一个谎言);新冠病毒应该就是Ralph
S. Baric等人在Defuse项目实施过程中,或Ralph S.
Baric自行尝试Defuse项目有关设计时制造出来的(稍后将说明Ralph S. Baric很可能是Defuse计划书的主要执笔人)。
IV 2018年1月生态健康联盟向Baric支付秘密酬金
据U.S. RIGHT TO KNOW(美国有权知道)网站披露:
EcoHealth Alliance paid Dr. Baric an undisclosed sum as honorarium (January 2018).
(生态健康联盟2018年1月向巴里克博士提供了一笔未公开的酬金)
https://usrtk.org/tag/ecohealth-alliance/
https://usrtk.org/wp-content/uploads/2020/12/EHAfunds_2017_Baric-Files.pdf

生态健康联盟行政助理要求巴里克为所收酬金填写W-9税务表格的Email
生态健康联盟因何向Baric支付酬金?可能有两个原因:
1、预支给巴里克的Defuse项目研究资金或项目启动资金;
2、巴里克是Defuse计划书的主要执笔人,或巴里克与达萨克是计划书的联合执笔人,这是巴里克撰写计划书的酬金。
以下时间线可能包含着新冠病毒产生,新冠疫情来龙去脉的一些重要线索:
1、2017年12月19日,川普共和党政府撤销了奥巴马禁令,全面重启了危险病原体的功能增益研究(恢复了举国性质的功能增益研究),重新为从事危险病原体功能增益改造研究的美国科学家提供联邦资金;
2、大约同一时期,美国国防部高级研究计划局(DARPA)启动了一项名为defuse的冠状病毒研究、改造计划,并将之委托给生态健康联盟组织实施;
3、2018年1月,生态健康联盟向Defuse项目的绝对技术核心巴里克支付了一笔秘密酬金,这可能是该项目的启动资金或撰写Defuse计划书的酬金;
4、2018年3月下旬,生态健康联盟将Defuse计划书提交至DARPA审核,并申请1400万美元的项目经费。Defuse项目包含多项冠状病毒功能增益改造课题,包括在冠状病毒中插入furin酶切位点,包括研究冠状病毒与宿主免疫系统的相互作用等等;
5、2019年7月5日,华裔病毒学家邱香果和他的丈夫,另一位华裔生物、病毒学家成克定,以及他们的若干中国学生被加拿大骑警强制带离加拿大国家微生物实验室(NML);
邱香果夫妇曾安排其学生将约15种高危、极高危活体病毒经由2019年3月31日多伦多-北京的加航航班寄送至北京。加拿大政府至今仍以“危及加拿大国家安全,危及国际关系”为由拒绝公开邱香果事件未经修改、遮盖的二百多页文件,加拿大政府对邱香果事件多次撒谎,并一直捂瞒病毒邮件的收件人信息;
邱香果本人是著名的免疫学专家和埃博拉病毒研究专家,其丈夫成克定是艾滋病研究专家。新冠病毒有极其全面的宿主免疫系统对抗机制;新冠病毒中存在与埃博拉病毒相似的结构(具有免疫屏蔽功能的O-Linked聚糖结构);新冠病毒中还有与艾滋病病毒高度相似的结构(gp120蛋白和Gag蛋白,gp120蛋白可帮助病毒感染CD4+T淋巴细胞,破坏免疫系统)。埃博拉病毒和艾滋病病毒都不是冠状病毒;
加拿大NML经常与美国合作开展病毒研究,其合作者包括美国国防部。邱香果与同事库宾格合作发明的著名埃博拉病毒治疗药物ZMapp中包含三个人源化抗体,这些抗体并非邱香果、库宾格、NML的原创,它们是十几年前美国军方资助项目的研究成果;
加拿大卫生局(PHAC)多次声称,加拿大NML定期跟其他公共卫生实验室共享病毒样本。
6、2019年11~12月,新冠病毒在武汉被发现。新冠病毒具有多项明显的人为设计特征,比如,它有同支系所有近亲都不具有的S1/S2处的furin酶切位点;再如,新冠病毒集免疫破坏、免疫屏蔽、免疫逃避、免疫抑制、免疫干扰等宿主免疫对抗机制于一身,在此之前,从未发现过任何一种冠状病毒,任何一种非冠状病毒具备如此全面的免疫对抗机制。
V Defuse 项目共同参与者王林发建议Baric回避反对“阴谋论”的科学界联署
2020年2月19日,国际顶级医学期刊《柳叶刀》(The Lancet)在线发表了一篇由8个国家(英、德、美、澳、荷兰、西班牙、马来西亚、中国香港)的27名著名病毒学家、流行病学家联合签署的一份声明:
Statement in support of the scientists, public health professionals, and medical professionals of China combatting COVID-19
支持抗击COVID-19 的中国科学家、公共卫生专业人员和医疗专业人员的声明
https://www.thelancet.com/lancet/article/S0140-6736(20)30418-9
声明中说:
We
stand together to strongly condemn conspiracy theories suggesting that
COVID-19 does not have a natural origin. Scientists from multiple
countries have published and analysed genomes of the causative agent,
severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), and they
overwhelmingly conclude that this coronavirus originated in wildlife, as
have so many other emerging pathogens. This is further supported by a
letter from the presidents of the US National Academies of Science,
Engineering, and Medicine and by the scientific communities they
represent. Conspiracy theories do nothing but create fear, rumours, and
prejudice that jeopardise our global collaboration in the fight against
this virus. We support the call from the Director-General of WHO to
promote scientific evidence and unity over misinformation and
conjecture.
我们站在一起,强烈谴责暗示COVID-19
并非自然起源的阴谋论。来自多个国家的科学家已经发表并分析了SARS-CoV-2的基因组,他们压倒性地得出结论,这种冠状病毒与许多其他新出现的病原体一样起源于野生动物。美国国家科学院、工程院和医学院院长以及他们所代表的科学界的一封信进一步支持了这一结论。阴谋论只会制造恐惧、谣言和偏见,危害我们在抗击这种病毒的全球合作。我们支持世卫组织总干事呼吁促进科学证据和团结,反对错误信息和猜想。
很多人以为联署者们是在为武汉病毒研究所、为中国说话;其实,这些科学权威、大腕们是在为科学界的领头羊,开展举国性质功能增益研究的国家—美国捂盖子。这份打着支持中国抗击疫情旗号的声明的真正目的,是第一时间围剿、扼杀“实验室来源论”,打着科学的旗号堵口科学辩论,消声科学质疑(合理质疑是最基本的科学精神)。
注:“实验室来源论”一直被捂盖集团阉割、偷换为“实验室泄漏论”,以回避、隐匿、掩盖武汉之外的实验室来源。
冠状病毒顶级国际权威Ralph S. Baric的名字并没有出现在联署名单中。
在这一科学界声明发表前两周,约在2020年2月5日,王林发给彼得·达萨克写信,建议他们三个人(王林发、达萨克、巴里克,都是Defuse项目成员)不参加科学界联署,并与之保持距离;
注:关于王林发的一些背景信息。
王林发是杜克-新加坡国立大学医学院新发传染病研究所所长,杜克-新加坡国立大学由美国杜克大学(Duke
University)和新加坡国立大学(National University of Singapore,
NUS)联合创办,于2005年4月成立。王林发还是武汉病毒研究所新发传染病中心科学顾问委员会主任。
王林发在Defuse项目中的分工是(参见前面章节III中的Defuse管理分工图):
Prof.Wang,
Duke-Nati,Univ.Singapore, will lead work on broadscale immune boosting,
building on his group's pioneering work on bat immunity.
杜克-新加坡国立大学医学院的王(林发)教授将领导广谱免疫增强工作(Baric在项目中领导靶向免疫增强工作),基于他的团队对蝙蝠免疫的开创性工作。
2月6日,达萨克将王林发的意见转达拉尔夫·巴里克,并用邮件标题强调:“No
need for you to sign the Statement
Ralph!!”,达萨克在邮件中还特意强调,他(达萨克)将设法使科学界的联署不会关联到他们三人的(病毒研究)合作。
巴里克当天复信,认同这一决定,并称,(我们参加联署)看起来是在自我服务,这有失(损害)(联署)的影响力。
巴里克和王林发二人依约没有参加联署,但达萨克经过斟酌,最终还是参加了联署(达萨克是联署的重要推动、促成者之一)。

达萨克向巴里克转达王林发提议不签署声明的邮件及巴里克的复信
上述邮件及回避科学界联署一事的合理解释是:
1、新冠病毒的设计、产生与美国国防部高级计划研究局(DARPA)启动的Defuse项目高度相关;
2、Defuse项目的三个重要成员巴里克、达萨克、王林发或者是新冠病毒的核心设计人,或者是新冠病毒设计、制造的知情人,心中有鬼的三人刻意回避科学界谴责“阴谋论”
的2.18联署,是不愿意世人联系、关注、了解他们参加Defuse冠状病毒改造项目情况;
3、新冠病毒的核心设计者,就是在Defuse项目中分工负责对蝙蝠冠状病毒实施功能增益改造,负责插入furin酶切位点,负责领导靶向免疫增强工作的拉尔夫·巴里克。
(未完待续)
| 

