新闻简述:9月3日著名医学期刊《医学病毒学》(Journal of Medical Virology)发表了数十位来自世界各地的科学家联名致信该期刊主编的经同行评议过的信件。该文对新型冠状病毒来源于蝙蝠冠状病毒,特别是RaTG13的行业共识提出了质疑。新型冠状病毒是第七种人冠状病毒,但也是第一个且是唯一一个具备大流行潜力的人冠状病毒。然而,其宿主取向/适应模式与其它冠状病毒存在显著差异,这就引起了关于新型冠状病毒的近端起源的讨论。
Questions concerning the proximal origin of SARS-CoV-2
关于新型冠状病毒近端起源的问题
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1002/jmv.26478.
Keywords: Coronavirus, Genetic Variability, Mutation, Fusion Protein
关键词 :冠状病毒,遗传变异,突变,融合蛋白
Summary:
总结:
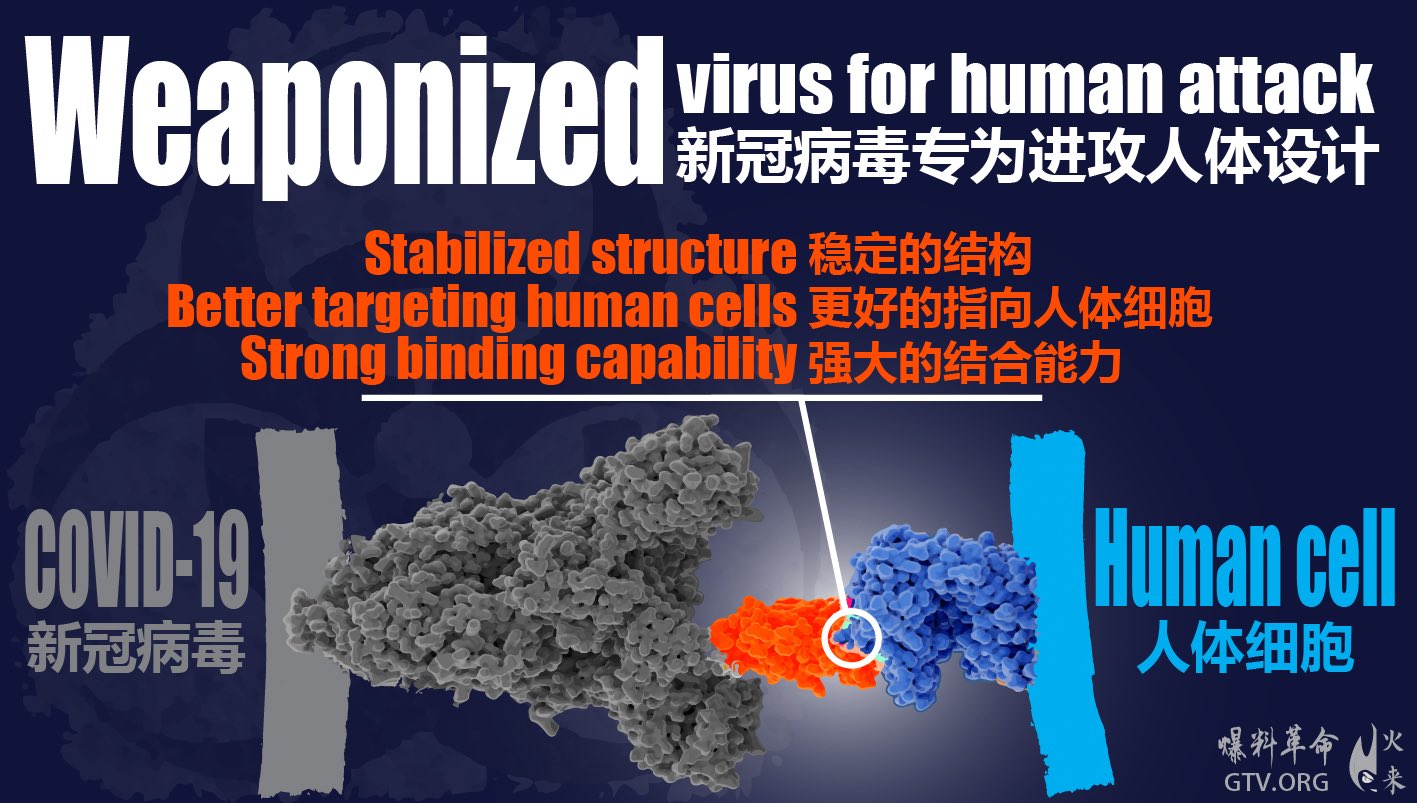
There is a consensus that Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) originated naturally from Bat coronaviruses (CoVs), in particular RaTG13. However, the SARS-CoV-2 host tropism/adaptation pattern has significant discrepancies compared to other CoVs, raising questions concerning the proximal origin of SARS-CoV-2. The flat and non-sunken surface of the sialic acid-binding domain of SARS-CoV-2 spike protein (S protein) conflicts with the general adaptation and survival pattern observed for all other CoVs. Unlike RaTG13, SARS-CoV-2 recombination presumably occurred between the S1/S2 domains of S protein enabling host furin protease utilization. Although millions of recorded cases have been recorded globally, SARS-CoV-2 S protein does not have any apparent further recombination, placing it in conflict with the recombination models of other CoVs. Similarly, the S protein receptor-binding domain (RBD) of SARS-CoV-2 has not accumulated high-frequency non-synonymous substitutions, differentiating SARS-CoV-2 from other CoVs that have positive selection/adaptation mutations in their RBDs.
Andersen and colleagues documented the possible natural origin of SARS-CoV-2 from BatCoV RaTG13 1,2. SARS-CoV-2 is the seventh zoonotic CoV virus capable of infecting humans, but the first and only human coronavirus (HCoV) with pandemic potential 3. Bat or rodent CoVs demonstrate certain specific changes in the S protein receptor-binding domain (RBD), as well as the S protein glycan-binding N-terminal domain (NTD), during host tropism/adaptation 4,5. SARS-CoV-2, unlike other CoVs, does not have those signature changes, suggesting that these RBD and NTD subdomains are of very recent origin.
The “Canyon Hypothesis” explains the development of canyons, depression zones or cavities on the surfaces of influenza virus, human rhinovirus, and Meningo viruses 6 . In CoVs (except SARS-CoV-2), the S protein NTD domain has several predicted glycanbinding domains, with a common feature being the hidden localization of these glycan binding domains to cavities to limit their access to antibodies and immune cells 5 . This pattern of CoVs is thought to be an evolutionary measure to restrict the recognition of these active sites by host immune system 4 .
HCoVs can evade detection by host glycan-binding immune receptors. Comparative genomic analysis of six HCoVs with their corresponding native bat or rodent CoVs suggests compatibility with the “Canyon Hypothesis” resulting from various adaptive S protein NTD non-synonymous mutations near or at the glycan binding domain which are predicted to result in these NTD domains being hidden below the protein surface5 . The predicted flat, non-sunken pattern of the SARS-CoV-2 S protein NTD glycan binding domains conflicts with this evolutionary host tropism/adaptation strategy 7.
A template-switching mechanism is presumably responsible for the high rate of RNA recombination in CoVs. In host cells, CoV RNAs show discontinuous RNA synthesis materialized by pauses of the RNA-dependent complex and subsequent jumps to downstream template acceptor sequences. This process results in subgenomic minus strand RNAs which serve as templates for subgenomic mRNAs. Due to the mechanistic similarity to recombination, this process might be at the origin of recombinant CoVs co-opting other CoV or even host related sequences8. Instances include the mouse hepatitis coronavirus S protein NTD sialic acid binding domain, likely arising from recombination of viral RNA with human galectin RNA sequences 8.
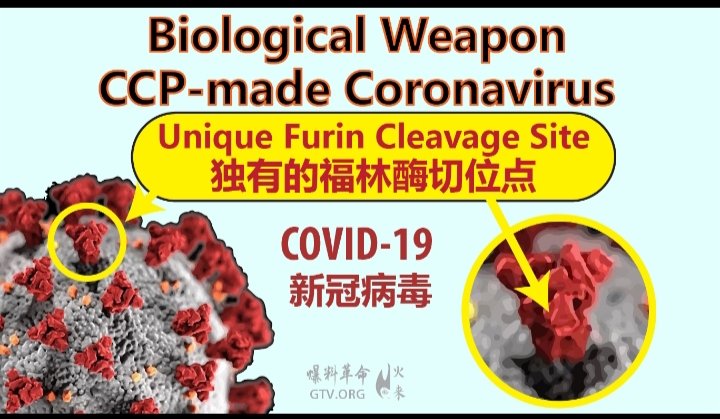
The furin recognition motif present at the SARS-CoV2 S1/S2 junction has no analogy in other “linage B” beta-coronaviruses, including neither pangolin-CoV nor RaTG13 1 . This indicates that the S protein S1/S2 junction is not a hot spot for RNA recombination termination that depends on a pattern swapping templates (copy-choice) 8 . Additionally, clinical isolates of SARS-CoV-2 S protein have not indicated any further recombination in this S1/S2 area, suggesting that the addition of a motif for S1/S2 site furin cleavage constituted a unique recombination occurrence. Finally, the CoV-unique insertion of 4 amino-acids creating a novel RRAR furin cleavage site introduces two arginine codons CGG-CGG, whose usage is extremely rare in CoVs, further supporting the hypothesis of a unique recombination occurrence.
HCoVs have high-frequency “hot spots” for non-synonymous amino acid replacements that can possibly create positive selection for host tropism/adaptation, resistance to neutralizing antibodies, or immune evasion 2 . Interestingly, clinical SARS-CoV-2 isolates to date have only a single high frequency non-synonymous mutation, D614G, in their S protein 9. Based on currently known mutation rates and patterns in clinical isolates of SARS-CoV-2, the S protein does not appear to be a mutational “hot spot” for SARSCoV-2, unlike other human CoVs.
SARS-CoV-2 is the seventh HCoV, but the first HCoV with pandemic potential. SARSCoV disappeared without a pandemic, and MERS-CoV is mostly endemic to the Arabian Peninsula with some additional limited traveler infections resulting in outbreaks in South Korea 3,4. These unique features of SARS-CoV-2 raise several questions concerning the proximal origin of the virus that require further discussion.
A. M. B, V. N. U, A. L., and K. L. provided critical review.
N. R., D. P., A. L. S. P. S.. V. N. U, K. L.T. M. A. edited the article,
M. S., P. A., T. M. A. formatted the article.
All authors M. S., D. P., P. A., T. M.A., S. S. H., A. S., R. K., K. L., D. M., M. T., A. L., P. P. C., V. N. U., S. P. S., B. D. U., N. R., A. M. B. and G. Z. K. of the consortium interpreted the results and approved the final version for submission.
M. S. 构思该研究.
A. M. B, V. N. U, A. L., and K. L. 进行重要审稿
N. R., D. P., A. L. S. P. S.. V. N. U, K. L.T. M. A.撰写文章,
M. S., P. A., T. M. A. 排版该文章
所有作者(M. S., D. P., P. A., T. M.A., S. S. H., A. S., R. K., K. L., D. M., M. T., A. L., P. P. C., V. N. U., S. P. S., B. D. U., N. R., A. M. B. and G. Z. K. )联合解释该结果并同意提交最终版本。
Competing Interests statement: The authors declare no competing interests.
利益冲突声明:作者声明没有利益冲突。
References:
参考文献:
1. Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF. The proximal origin of SARS-CoV-2.NatMed.2020;26(4):450-452.doi:10.1038/s41591-020-0820-9;
2. Malaiyan J, Arumugam S, Mohan K, Gomathi Radhakrishnan G. An update on the
origin of SARS-CoV-2: Despite closest identity, bat (RaTG13) and pangolin derived
coronaviruses varied in the critical binding site and O-linked glycan residues [published online ahead of print, 2020 Jul 7]. J Med Virol. 2020;10.1002/jmv.26261. doi:10.1002/jmv.26261
3. Forni D, Cagliani R, Clerici M, Sironi M. Molecular Evolution of Human Coronavirus Genomes.Trends Microbiol. 2017;25(1):35-48.
doi:10.1016/j.tim.2016.09.001
4. Hulswit RJ, de Haan CA, Bosch BJ. Coronavirus Spike Protein and Tropism Changes. Adv Virus Res. 2016;96:29-57. doi:10.1016/bs.aivir.2016.08.004
5. Li F. Receptor recognition mechanisms of coronaviruses: a decade of structural
6. Rossmann MG. The canyon hypothesis. Hiding the host cell receptor attachment
site on a viral surface from immune surveillance. J Biol Chem.1989;264(25):14587- 14590.
7. Fantini J, Di Scala C, Chahinian H, Yahi N. Structural and molecular modelling
studies reveal a new mechanism of action of chloroquine and hydroxychloroquine
against SARS-CoV-2 infection. Int J Antimicrob Agents. 2020;55(5):105960.
doi:10.1016/j.ijantimicag.2020.105960
8. Makino S, Keck JG, Stohlman SA, Lai MM. High-frequency RNA recombination of murine coronaviruses. J Virol. 1986;57(3):729-737. doi:10.1128/JVI.57.3.729-737.1986
9. Brufsky A. Distinct viral clades of SARS-CoV-2: Implications for modeling of viral spread [published online ahead of print, 2020 Apr 20]. J Med Virol.