Up to date, many reports have described the association between respiratory viral infections with neurological symptoms. There are several recognized respiratory pathogens that gain access to the central nervous system (CNS), for instance, respiratory syncytial virus, the influenza virus, the human metapneumovirus, and HCoVs (HCoV229E, HCoV-OC43, and SARS-CoV) [39], that induce manifestations such as febrile or afebrile seizures, among other encephalopathies [40, 41].
Primary cultures of human astrocytes and microglia and various human neuronal cell lines, such as the neuroblastoma SK-N-SH, the neuroglioma H4, and the oligodendrocytic MO3.13, have potential tropism for HCoV-OC43 [42]. Using an experimental animal model, HCoV-OC-43 infection also showed neuro-invasiveness and neuro-virulence [43]. Therefore, it is not surprising to find brain SARS-positive autopsies. Using in situ hybridization, the SARS genomic sequence has been detected in the cytoplasm of neurons of the hypothalamus and cerebral cortex [44]. Furthermore, Moriguchi et al. [45] confirmed the presence of the new SARS-CoV-2 in cerebral spinal fluid. In accord, epidemiological and clinical research have described neurological, non-common symptoms, and neurological manifestations associated with the SARS-CoV-2 infection (Table 2). These clinical features include neuralgia, confusion, hyposmia, hypogeusia, and altered consciousness, symptoms that evidence the neurotropic invasion by SARS-CoV-2 [19, 41].
Table 2 Neurological manifestations associated with SARS-CoV-2 infection
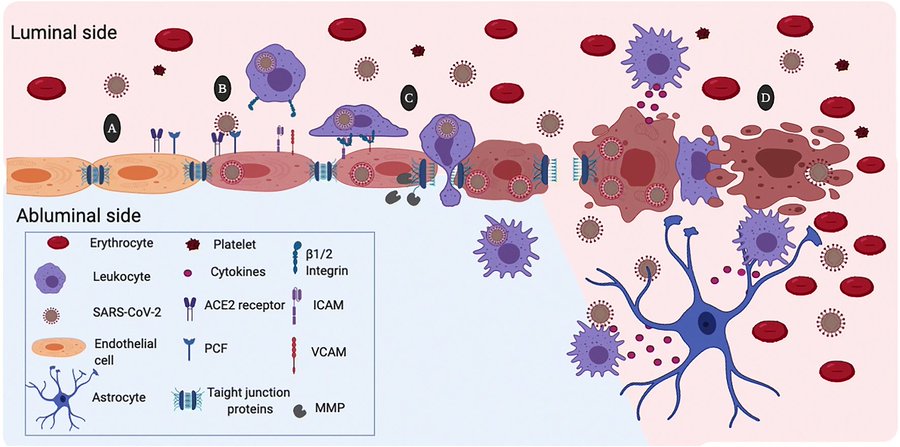
Despite the evidence demonstrating the neurotropism of respiratory viruses, the exact mechanism of neuro-invasion accomplished by viruses remains currently unknown. However, the route of invasion of the CNS has recently been described for HCoV-OC-43. This virus gains access to the CNS through the olfactory bulb, moving along the olfactory nerve. Then, neuro-propagation occurs along the multiple axonal connections expanding through the CNS (e.g., neuron-to-neuron propagation or diffusing particles) [56]. Similar to HCoV-OC-43, a model in vivo of SARS-CoV infection suggested that the virus enters the brain via the olfactory bulb, and then, a transneuronal spread could occurs [57].
Also, some infectious blood-borne viruses primarily targeting peripheral organs have evolved strategies to thwart the blood-brain barrier (BBB). These strategies include direct infection of the brain microvascular endothelial cells that form the BBB, a paracellular entry that involves alteration of the tight junctions, or the “Trojan horse” invasion, via the traffic of infected monocytes/macrophages migrating across the BBB, in a similar manner as the not-respiratory immunodeficiency virus 1 (HIV-1) [58, 59]. Likewise, when human primary monocytes are activated following infection by HCoV-229E and eventually become macrophages, it can invade tissues, including the CNS [60, 61]. Additionally, it has been reported that through activation of the brain microendothelium, the damage caused by the inflammatory response, allows the virus to reach the CNS. In this sense, the neuro-invasion of SARS-CoV-2 could occur through trans-synaptic transfer, via the olfactory nerve, infection of vascular endothelium, or leukocyte migration across the BBB [38].
Recent information has shown the SARS-CoV-2 ability to infect CNS cells, especially the brain microvascular endothelial cells of the BBB. This situation explains the neurological symptoms observed during infection and reveals the possible consequences of viral infection. Although it is too early to elucidate the long-term side effects of SARS-CoV-2 infection, the background obtained with other respiratory viruses suggests that SARS-CoV-2 might induce permanent sequelae in the CNS through damage to the BBB, including dementia in predisposed patients. Furthermore, the proinflammatory state, due to viral infection seems to be the general mechanism involved in the induction of BBB damage. However, further studies are necessary to confirm this evidence.