我们是由北京猿人进化而来的么?
答案可能是NO.
我们是多民族融合而成的么?
答案可能还是NO.
我们是谁,我们从何而来?
2003年4月14日,美国科学家在华盛顿庄严宣布:美、英、日、法、德和中国,6个国家联合,经过13年的努力,共同绘制完成了《人类基因序列图》。由IBM赞助,美国《国家地理》杂志举行的研究“人类迁徙遗传地理图谱计划”2005年4月开始在世界各地进行,复旦大学生命科学院承担了远东地区及其东南亚地区的DNA的取样和研究。
随着研究的深入,答案渐渐浮出水面,一张完整的人类迁移路线图开始显现。我们整理了一些大家比较关注的内容。
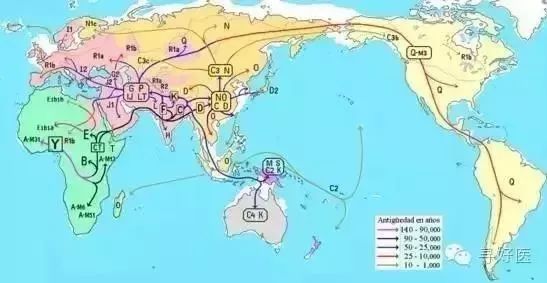
全世界人口基因来自36个女人
源于“线粒体夏娃”
令人意想不到的是我们60多亿人口可能源自一个母亲。英国牛津大学人类遗传学家经十几年的DNA研究发现,全世界的人口分别繁衍自36个不同的、被称做“宗族母亲”的原始女人,其中非洲有13个“宗族母亲”,印度、澳大利亚、中亚有7个。而所有这些“宗族母亲”又都是15万年前到20万年前非洲大陆上一个科学家命名为“线粒体夏娃”(Mitochondrial Eve)的女人的后代。尽管“夏娃”不是当时惟一活着的女性,然而她却是惟一一个将血脉延续繁衍到今天的原始女人。
据英国《每日快报》报道,该理论是由英国牛津大学人类遗传学教授布赖恩·西基斯(Bryan Sykes)提出的,他是世界上第一个发明从年代久远的古代骨骼中提取出DNA方法的遗传学者,并建立了“牛津祖先”(Oxford Ancestors)项目。
西基斯教授研究发现,现代欧洲人其实大多数都是远亲:97%的现代欧洲人,其实都起源于10000年前到45000年前冰河时代的7个不同女人,这7个“宗族母亲”被他称做是“夏娃的7个女儿”,7名原始女人通过“线粒体DNA”和现代欧洲人联系到了一起。线粒体DNA是只通过母系一脉传递的遗传基因,男性也能从母亲那里继承线粒体DNA,但却无法将它遗传给自己的后代。也就是说,如果一个女性生下的全都是儿子,那么她的线粒体DNA遗传链将因此终止。所以,如果一个女人没生出女儿,那她就真的断子绝孙了。
西基斯称,“很显然,在漫长的冰河时代,欧洲并非只生存着这7个女人,同一时代生活着大量的原始女性。然而她们要么没有长到成年,要么没有生下孩子,要么生下的全是男孩,但这7个原始女人不仅活了足够长时间,并且每人至少生了两名女儿,从而开始了线粒体DNA遗传链,并且一直延续到了今天。 据西基斯教授称,线粒体DNA一般很难发生改变,平均要过2万年线粒体DNA才会发生微小的变异。他对记者道:“研究发现,大多数现代欧洲人的线粒体DNA可分为7种类型,就仿佛血型一样。我们相信,每个线粒体DNA相同的人都是数万年前同一个女人的后代。”
西基斯承认,研究人员也偶尔发现有的人DNA不属于任何一个宗族母亲。他开玩笑地说,或许来自外星人吧。” 如果你想知道自己的“宗族母亲”是这36位中的哪一个,180英镑就可以搞定,上网联系“牛津祖先” www.oxfordancestors.com。已有二万人在“牛津祖先”的帮助下认祖归宗。
民族性只是社会概念
其实人类无种族之分
我们在广播电视里会听到民族问题,但是也许这个只是一个社会概念,并不是一个科学概念。
美国最早宣布破译人类基因密码的塞莱拉公司负责人万特博士说,基因图谱并未显示‘种族’之间有何差异。我们都是10万年前从非洲的少数原始部落迁移和进化而来。人类只有一个种族。
大多数科学家开始相信,区别人类种族的那些标准实际上与生物学概念几乎没有什么直接的关系。他们说,区别不同种族最常用的特征,如皮肤和眼睛的颜色、鼻子的宽度等,是由相对而言极少数的基因控制。10万年时间只是历史长河中的片刻,在这短暂的人类进化史上,这些基因的变化是为了适应环境所带来的巨大压力,例如,赤道地区的人皮肤黝黑,是为了减少紫外线辐射的伤害;而北方地区的人皮肤白皙,是为了在较弱阳光下人体能产生较多的维他命D。
纽约曼哈顿北区总医院院长佛理曼说:“如果你一定要问人的外表不同反映出多少基因组的差异,我的回答是:不到0.01%,这只占基因总图谱极小极小的一部分。”亚特兰大艾摩利大学医学院分子遗传学教授华莱士说:“不幸的是,人类特别关注外表细节的差异,夸大所谓‘种族’差别的意义,似乎我们整个社会的结构都建筑在视觉的基础上。”
科学家们认为,与极少量基因决定人的肤色和外表相反,人的智力、艺术天赋和社交能力等却由人类8万个基因中数千甚至数万个基因所决定,而且是以复杂而相互关联的方式起作用。科学家们还发现,生活在同一地区的人,某方面基因的差别之大可达90%,而因生活地区不同而产生的基因差别只占10%。有些基因,例如控制免疫系统的基因,在人与人之间差别极大,可是这种差别与种族没有任何关系。
加拿大安大略省的印度裔生物学博士安南德建议,在研究人种时,与其去考虑种族不同,不如从临床的角度去寻找某一种群体易染上何种疾病,例如印度人形成血栓的几率较高,这也许是因为印度的文化和人们的生活习惯增加了引起心脏疾病的危险。所以,在考虑人种时,应从生物学的角度,全面考虑他们的生活方式、饮食习惯等,而不是强调他属于哪个种族。
但也有少数生物学家对此持有异议。犹他大学人口遗传学家罗格斯就认为,种族划分是有意义的。他说:“我们也许相信种族之间的区别主要表现在表面上,但差异确实存在。种族分类对于研究人类的起源和迁移过程有很大帮助。”
我们的祖先如何来到中国
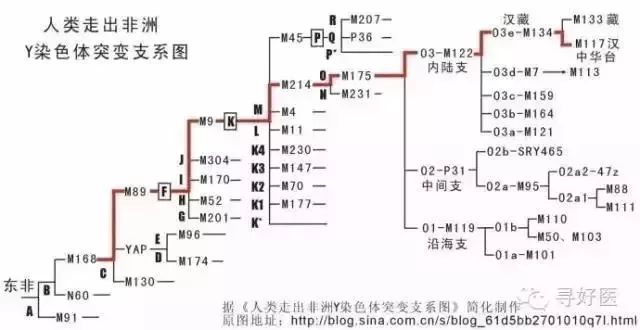
科学家意想不到的是汉人和藏人本出同源,分子人类学家根据研究DNA得到了这一结论。而且,汉人与藏人的关系比苗人、越人的关系都要更接近。汉人和藏人的分开不过是5000年前的事情。

中科院昆明动物研究所研究员宿兵于1996年在DNA中找到了汉藏同源的证据。他发现汉人和藏人在M122以及在其分支M134上都有相同的突变。而且宿兵通过对Y染色体主成分进行分析,发现藏族的突变频率与汉人最接近,这意味着在藏缅语系的诸多民族中,藏族和汉族最接近。他们分化的年代约在5000年前。
M122突变大约发生在三四万年前,那时中国陆地上的许多山脉被积雪常年覆盖。棕色人种早于黄色人种在黄河、长江流域活动,在东南亚的缅甸一带,黄种人的部落已经取得一定势力,棕色人不断沿海岸线退缩,或者躲进山区。“当中国陆地上的冰川不断消融时,一支带着M122突变的南亚语人群开始进入了中国。”李辉说,目前发现关于南亚语先民进入中国后的分化路线有3条,共有两个入口,一个是在云南,一个是珠江流域。
其中一支沿着云贵高原西侧向北跋涉,他们最终在距今1万年前的时候到达了河套地区,黄河中上游的盆地。“这里应该是中华文明的真正起源地。”汉藏语系的祖先也被后人称之为先羌,他们也就是汉族与藏族人的祖先。
这一支南亚人在出发时头颅还很圆,带有大鼻子、厚嘴唇等特征。但在他们行进过程中,人体形态发生变化。而到了高原之后,不再受疟疾影响,加上缺氧,存活下来的人基因变化了,脸都变得很长,线条、棱角变得刚硬,如同刀削。留在河谷地区的羌人应该是迁移过程中留下的。他们保留了游牧的习性。
经过这1万多年的迁徙,这样一直到5000~6000年前,人体内部的DNA又开始不安分了,那时,由于粟谷农业的出现,新石器文化开始在这个地区发展。人口的增长使群体必须扩增新的居住地。汉藏语系的两个语族开始分野。
其中一个亚群在M134的基础上又发生了M117的突变(汉族特有的两个基因突变)。他们带着这个突变向东行走,一直到渭河流域才停留下来。他们掌握了农业文明,开始以农耕为生。这个群体就是华族,也就是后来所称的汉人的前身。李辉在复旦大学生命科学学院攻读博士学位。研究方向是分子人类学,他热衷收集中国及东南亚地区各个民族分支的DNA样本,足迹遍及中国南部的城镇与村落。“M117是汉人身上很古老的一个遗传突变。”李辉说。
另一个亚群被称为藏缅语族群体。他们或是战败、或是寻找食物离开黄河流域,向西向南迁移,最后在喜马拉雅山脉南北居住下来。(藏族)在迁徙的过程中,这个群体就像细胞在不断裂变,分化出了藏、彝、景颇。至今,这些民族的送魂歌中都要把亡灵送回北方。“藏族在形成中吸取了中亚的许多因素。”
华夏族扩张很厉害,他们向四面掠夺土地和粮食,羌族这时候不再被他们认做是同类,称其为“西戎”,当然也包括东部的广泛地区,但这一地区已经被“三苗”占领,这是和汉藏同一祖先而更早迁移到此的族类,部族的争斗产生了民族英雄,“黄帝”就是这样的人物。而后,华族和东夷、南蛮在夏商阶段一直在争斗中。“楚人是荆蛮的后代,从荆蛮里面分化出三苗,所以,现在分子遗传学可以看出,很多湖北人DNA和苗人的关系接近,胜过与汉人的关系。”李辉说。
第二条路线是珠江流域。百越先于汉藏进入两广地区,穿越南岭的变为北越,也就是后来越国的祖先,南方的越人自秦汉以来一直往广西山区迁移,南宋时候又发生迁移,后来形成侗族。所以,现在浙江一带越人的语言和侗族语言有相同之处。而吴国人则有不同的起源,吴越两国根本不同源,统一使用华语后才接近。历史学的猜测再次失败。
吴国人从何而来呢,当初南亚先民从云贵高原西侧进入中国后,一支人群与汉藏民族分开而行,他们沿着长江往下走,这个人群在洞庭一带形成了苗瑶语系。“吴人就是苗瑶人东进和南下的汉人同化后形成的,所以他们遗传结构的变化多而复杂。”
在渭河流域华夏族逐渐磨砺出一支骁勇善战的“”秦人“”,他们盘踞在关中,积聚着统一中国的实力,秦汉时期,四分五裂的中国被统一,从语言、文字到货币、计量单位等文化层面上,汉族的整体意识真正形成。加上强有力的中央集权统治,之后汉族就没有再出现分化。而且,各地的汉族基本没有带上外来的成分。他们通过先进生产力而不断地扩张,占据了大部分土地。“历史学家说的汉人扩张中不断融合当地人是不对的,当地人基本被驱逐了,福建人是最好的证明根据分子人类学分析,他们基本上是北方汉人的后代,语言上保留古代汉语尤其多。”李辉说。
金力教授的实验室助理钱吉说,其实中国在利用DNA分析自己祖先的迁徙路线上已经相对比较成熟了,就目前的研究成果来看,华夏56个民族和东亚东南亚各民族都是由南亚语系的先民分化出来的,因为他们身上都带有M122的突变。
人类基因图谱
——世界人口基因组分布
1987年,美国夏威夷大学的瑞贝卡·坎恩破译了来自世界各地的妇女的线粒体DNA,发现现代女性的线粒体DNA都来自一位妇女,她大约生活在15万年-20万年前的非洲。
随后,分子人类学家再次成功破译了男性遗传密码的Y染色体。通过研究,他们得出结论,现代男性都有一个共同的父亲,他生活的年代也应该在大约15万年前的东部非洲。
“DNA中的遗传密码让亚当、夏娃离开了神话世界,变成了有血有肉的非洲人。他们成了包括中国人在内的所有人的祖先”。复旦大学生命科学学院博士生李辉说。
李辉解释说。大约15万年前,在东非分化出了很多人种与部落,其中就已经包含了现在的黑、棕、黄、白四个人种的祖先。Y染色体上的M168是目前发现的一个很古老的突变位点,这是人类在要离开非洲时产生的突变,大约发生在10万年之前。那些棕色人、黄种人就是带着这个古老的突变开始向世界扩散。除了非洲以外的现代人都具有这个位点的突变。

10万年前,地球处于冰川期,大部分陆地被冰川覆盖。整个海平面比现在低120米左右,许多海床裸露在地面。在东非,各个部落拥挤在这块炙热的土地上,抢夺着有限的食物。或许是频繁发生类似于现在的卢旺达种族清洗事件,一部分人开始走出非洲。
“又过了几千几万年,现代人在Y染色体基因突变M168基础上又出现了2个突变类型,M130和M89.前者发生在8万年前,后者发生在4万5千年前,人类已通过北非进入了欧亚大陆。”李辉说,最早出来的是棕色人。他们主要是在海边捕鱼为生,并捡食滩涂上的高蛋白食物。他们拿着旧石器和尖木叉,在印度洋岸边走着。公元5万年前,他们已占领了南亚和东南亚的陆地和岛屿。在之后的几万年里,散漫的捕捞捡食生活让他们沿着无边的海岸线,一路走到东亚,之后可能穿过白令海峡到达北美和南美洲。
黄种人走出非洲的时间要比棕色人晚了近5万年。但他们的扩散速度很快。他们只用了棕色人1/5的时间就到达东南亚。柯越海取样的1万2千个中国人的样本中有11311个样本在M89的位点上发生突变。而这个突变标记在黄种人到达东南亚时就已形成,并且一直在这里做了很长久的活动,之后他们将要进入中国,分化出现在的汉族。
我们再来看4个基因
D-YAP 矮黑人基因
C-M130 棕种人基因
K-M9 是黄种人基因
P-M45 是白种人基因
D-YAP矮黑人基因、M130棕种人基因,汉族人无论是南方汉人还是北方汉人,Y染色体中都没有这两种基因。
而北方民族蒙古人,突厥人,满族人,朝鲜人和日本人都有D-YAP和M130。
其中K-M9是黄种人基因,是中国人的主体类型。P-M45是白种人基因。
C—M130是棕种人基因,海洋居民,C—M130走的路线大概就是沿着海岸出发,从阿拉伯半岛—伊朗—印度—中南半岛,此后,这批C—M130的子孙分为两支系,NO.1支系向北进入西伯利亚,并最终进入了北美地区;NO.2支系向南进入澳大利亚,并扩散到整个太平洋诸岛。
今天C—M130的后代主要分布在印度南部、北亚东部、日本、北美西部、澳大利亚、太平洋诸岛。在东亚地区,外蒙地区比例非常高,达到58%,还有哈萨克人、吉尔吉斯人、布里亚特人、比例都在60%以上,这些人群的类型(包括北美地区)都是C3—M217;M130的其他类型还有,日本的阿伊努人90%以上是C1—M8,另外,现代日本人(6.5%)、朝鲜人、满州人、阿穆尔人中,也有一定数量的C1。C2—M38,主要分布在澳大利亚和太平洋诸岛屿,C4、C5主要分布在南印度地区,在当地民族中能达到20%。在汉族人中几乎没有C-M130基因的分布。
D—YAP又被称为小黑矮人基因,是一支非常古老的基因,它和C—M130几乎同时走出非洲。现在D—YAP类型主要分布在印度安答曼群岛的安达曼人(100%)、藏族(58%)、土家族、彝族、瑶族、日本(34.7%)、朝鲜、满州人、缅甸人、克钦人。在汉族人中,D—YAP在汉族人群中几乎没有发现。Y染色体变异数量多少并不能用人口数量来衡量,很多人口众多的大民族都是由一个小人群由于某种原因迅速壮大而来的,如历史上曾建立了强大帝国(典型的是汉人、阿拉伯人),近代的高技术革命和殖民(如英格兰人、西班牙人),这些人群的同源性强,Y染色体的变异类型并不特别丰富,相反,一些非常古老的小民族,其Y染色体变异类型却十分丰富。
根据分子生物学的研究,原来的汉族融合而成的理论是错误的,汉族的基因构成恰恰是一个非常单一纯粹的民族,K—M9基因是最重要的一个支系,亚欧大陆上,除了西亚地区和西伯利亚,世界岛上绝大部分民族都由K—M9的子孙构成,今天汉族人中,96%的人都是K—M9类型。(看来我们和日本人和朝鲜人还是有很大的不同)
K—M9下面也有很多子类型,最重要的有两个,一个是ON—M214,另一个是P—M45。前者(ON—M214)主要分布在东亚、东南亚、北亚东部、部分太平洋岛屿(以上都是O—M175子类型)和乌拉尔山两侧、北欧北部、东欧北部和北极圈内的爱斯基摩人(以上都是N—M231子类型)。中国人主体类型就是O—M175。族人中的4%的P-M45白种人基因,最有可能的来源的是古代中原的高加索人种。
北京猿人不是我们祖先
“我们的祖先究竟是谁?”
初中教科书的权威教育是,分布在中国的现代人是由生活在40万年前的北京猿人进化而来。这个说法的理论依据是人类的多地区起源说,认为在欧、亚、非的现代人都是由当地的猿人进化而来。
金力教授的实验室助理钱吉说,现代人是单一的物种,而各地的猿人差异却很大,属于不同的物种,并没有都进化成为现代人。只有东非的猿人走上了向现代人进化的道路。所以,北京古猿人不可能成为我们的祖先。
中科院昆明动物研究所研究员宿兵在查阅中国现有化石的年代以后,发现了一个不容忽视的断层。这个断层从大约10万年前至4万年前,没有任何人类化石出土。经推测,生活于东亚的直立人和早期智人(HomoSapiens)在最近一次的冰川时期,由于恶劣的气候而绝灭。取而代之的是从非洲不远万里迁徙而来的现代人种。
但仍有科学家支持“多地区起源说”。中科院古脊椎古人类所的吴新智院士细心比较北京猿人、山顶洞人和现代人的骨骼特征,发现70%的中国人在头骨上有3个特征和北京猿人是一致的。所以他依旧觉得北京猿人可能是中国人的祖先。但他的学生刘武去非洲考察时,却发现有30%的东非人在这3个头骨特征上也和北京猿人相一致。
1998年,中国科学家意识到DNA遗传密码的重要性。在那些双螺旋的DNA链条上交织着红、黄、蓝、绿4种颜色的小球,A、T、C、G.它们掌管着整个人类的迁徙和发展史。
按科学上的解释,每一个小球就是一个核苷酸单位。所谓基因就是这些小球按照一定秩序连在一起,并具有相应的遗传信息。它有很强的功能性,控制着人类的肤色、形态和健康。比如人类的高血压就有几百个基因在控制。它们的序列不能随便更换。变换了,就要影响身体状况。
而还有些小球的序列不具有功能性,性质如同DNA分子里的填充物。它们的排列是自由的。它们过了几千几百年会自由地发生一次变化,例如原本A的地方出现C.这种变化不受任何外界因素影响,不承担自然选择的压力。它们的改变不会影响身体健康。就是这些遗传突变记载了人类迁徙的历史信息。
中科院昆明动物研究所研究员褚嘉佑着重研究的是DNA排列中的一种被称为微卫星的特殊排列顺序,这是一种公认的遗传标记。DNA排列中有一种简短的重复,比如TGTGT鄄GTG,其重复的字母和次数都可以遗传,当延续到后代身上时,原先那一个G的位置可能会变成A,或者原先重复9次而到了后代忽然变成重复10次。根据这个原理,褚嘉佑分析了28个东亚人群,推算出在中国的现代人起源时间不会早于5万年。
有人认为褚嘉佑的取样数量太小,没有说服力。2001年,金力的学生柯越海和他的研究小组对主要以华人为主的东亚人群进行了大规模的遗传分析。他进入的是Y染色体的世界。它们一代代地由父子相传,而且“性格”稳定,一般在几十代之后才会有一到两个基因位点发生变化。这些遗传突变位点的结构如同一棵树,也正是这棵基因树记录了人类在不同时间向地球不同地点散布的路径。
柯越海共分析12127个男性个体,发现那些Y染色体的YAP、M130和M89上都会有其中一个位点发生突变。它们是基因树中的三根树枝。它们汇拢到一根叫M168的树根上。这个M168就是非洲人体内的突变位点。也就是说中国人与生活在东非的非洲人有关。
柯越海说,中国人Y染色体的有效群体本来就不大,所存在的种类不多,这1万2千多个样本几乎囊括了绝大部分中国人染色体的类型。这可以充分证明,华人占大多数的东亚人群起源于非洲。



