進化理論的數學模型 前幾天讀到胡大偉先生的一篇博文,文章不長,貼在下面。 進化很好理解,但怎麼用數學模型來表達呢?胡大偉 這些年,我一直在想下面的問題,並把它向很多外國大理論生態學家提出,可他們的回覆都是 I don't know。 Although it is very easy to intuitively understand Darwin's theory of evolution, i.e, ever increase complexity of system structure sustained by matter, energy and information connections for acclimation, I think that there is a more enormous challenge on how to express it by mathematical models whose analytical expression must also be changed along with time. For example, suppose that at t moment,the kinetic model of a specific ecosystem could be written as follows: dx/dt= f(t)(t,x,p,u) = 3*x*u- 2*exp(-x) wheret x, u is state and input, respectively. but at t+△t moment,it maybe changed by eco-work during △t into new analytical expression as follows: dx/dt= f(t+△t)(t,x,p,u) = x^2-2*exp(-2*x)+u*exp(x) If we can solve this huge problem, really valid ecological models would be developed to describe and predict the ecosystem behaviors, such as the species origin, community succession, niche differentiation, ecosystem evolution, and so forth. 這個問題不解決,近5年來,我所建立的生態學模型都很naive,因為和所有建模者一樣,把生態系統看成了機械系統,沒有觸及它的根本動力學機制。生態系統的變化不是量變,而是質變,我理解質變就是結構的複雜化。 ~~~~~~~~~~~~~~ 現在,生態系統的標準模型是常微分方程,這在他的博文中也可以看到。我在他的博文後面留言,“生命現象是熱力學現象,應該用熱力學方程描述。” 他回答,“我可以毫不猶豫地說,傳統的熱力學定律和方程根本無法描述生命運動。熱力學必須有最原始的創新,就像牛頓發明導數那樣的創新,才有可能把數學用在描述生命現象之中。” 他說得很對,早期的研究者都知道生命現象是熱力學現象,應該用熱力學方程描述。正是由於沒找到適當的熱力學方程,才用更簡單的常微分方程建模。我們從最原始的角度,看一下生命運動的根本特徵。生命運動的根本特徵應該包括兩條。首先,所有生物都需要吸收外部的資源來補償自身的耗散,第二,從外界得到的資源必須要高於成本,包括固定成本和可變成本。我們所說的自然選擇,加以量化,就是回報率大於零, 就是攝取的資源高於消耗的成本。下面我們將這兩條特徵轉化為數學理論。 這個理論的推導確實不容易,但是最終的結果是個解析解,非常簡單。理論推導出來後,我們用它來解釋 species origin, community succession, niche differentiation, ecosystem evolution, and so forth.
吸收資源以補償自身的耗散,這可以用對數正態過程來表達, 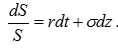 第一項是吸收項,第二項是耗散項。由於我們關注的是平均值,而非隨機分布的每一點,我們可以將隨機過程轉化為相對應的確定性方程。轉化方法有很多種,我們採用量子力學中的費曼路徑積分方法(費曼-Kac 公式),這是因為量子力學是最普遍的求加權平均的方法。
利用費曼-Kac 公式得到代表生命過程的熱力學方程是
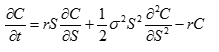
我們需要初始條件才能解這個方程。初始條件就是我們常說的自然選擇的量化。具體一點,我們把初始條件設成獲取的資源量等於成本,固定成本加可變成本,這是生命存在的邊界條件。如果固定成本低於所攝取的資源,可變成本是資源和固定成本之差,如果固定成本高於所攝取的資源,就不再需要額外的可變成本。讓 S 代表資源量, K 代表固定成本, C 代表可變成本,數學上,這就是
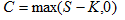
有了這個邊界條件,我們就可以解出這個方程,其中的時間值 T 代表生命的長度(lifespan), 或投資的期限。恰好可以得到如下的解析解, 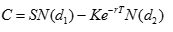 (1) (1) 其中 
而 N(x) 是標準正態分布隨機變量的累積概率函數。也就是說,可變成本可以表達成資源量,固定成本,生命長度或投資期限,耗散率或者不確定性,和折現率的解析函數。熟悉金融工程的讀者,會看到這個公式和 Black-Scholes 公式的形式一模一樣,但各個參數的經濟意義是不一樣的。 固定成本可以看成系統複雜性的量化標誌。 有了這個可變成本的解析公式,我們可以進一步計算各種投資的預期回報。假設整個投資期間的產出量是 Q,單位產品的價值和可變成本分別為 S 和 C, 則總收入和總成本分別 SQ 和 K + CQ
投資的淨現值為 SQ - (K+ CQ) (2)
投資的回報率為
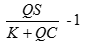 (3) (3)
我們經常用 S 代表單位時間的產出, 如果投資項目的期限是 T, 那麼, 投資的淨現值為 ST - (K+ CT) (4) 投資的回報率為 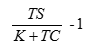 (5) (5) 有了這個生產過程的解析理論,就可以推演出經濟學和生態學中很多現象。我們先來計算在不同大小的不確定性環境下,固定成本增加時,對可變成本的影響。從公式 (1),計算結果如下圖。
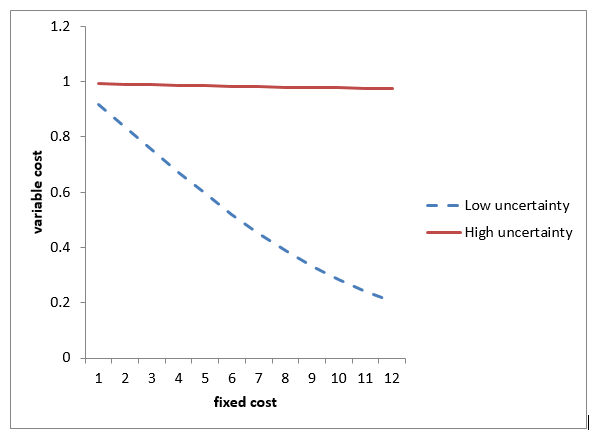 上圖的計算顯示, 高固定成本的投資在低不確定性的環境下能顯著降低可變成本, 而低固定成本的投資在高不確定性的環境下更具靈活性。在生態學裡,物種分類為低固定成本的r類物種和高固定成本的K類物種,在波動幅度很大的環境,r類物種 (r species) 有優勢,在穩定的環境裡,K 類物種 (K species) 占統治地位。從上面的計算,當耗散係數很大,代表環境變化很快,無論如何提高固定成本,很難降低可變成本,在這種環境下,低固定成本的r類物種占優勢。當環境穩定(耗散係數低),提高固定成本可以迅速降低可變成本,在這種環境下,高固定成本的K類物種占統治地位。 同樣,在變化很快的經濟領域,技術往往由一兩個人創造的新公司引領,象蘋果,微軟,網景,雅虎,谷歌,臉書,往往是幾個年輕人建立的,而成熟的領域,象消費品行業,往往是象可口可樂,寶潔這種老牌企業稱雄。在科學研究中,成熟的領域往往由名校的大牌教授領導,研究的規模龐大,而革命性的理論往往源於新人或局外人,由一兩個人完成。 這解釋了 niche differentiation。 我們再來計算不同固定成本的投資,在不同的產出或市場大小情況下的回報。從公式 (3),計算結果如下圖。 
從上圖可以看到,高固定成本的投資需要較大的產出才能回本,同時,由於高固定成本的投資可變成本低,其回報曲線較陡,在大市場中可以取得高回報。而低固定成本的投資則相反。
在一個新的生態環境中,比如說由於滑坡,環境營養缺乏,不穩定性高,從上圖可以看到,低固定成本的生命容易存活。當環境營養不斷積累,系統趨於穩定,高固定成本的生命更有競爭力,逐漸統治生態環境。
這解釋了 community succession, and ecosystem evolution. 不同的環境,需要不同的固定投資,這就是 species origin 的原因。
我們接下來討論人們在不同環境下的投資決定。在給定的條件下,人們會選擇特定的投資方式以得到最大的回報率或淨現值。我們假設不確定性是環境條件,是我們不能改變的,而固定成本和投資期限是我們自己可以調節,控制的。在 Excel 中,有一個 add-in, 叫 solver, 可以用來解決極值問題。在不同的不確定性下,我們以固定成本和投資期限為變量,用solver 來尋求淨現值,即公式 (4)的最大值,大家可以看到,當不確定性低的時候,固定成本高和投資期限長,淨現值高,當不確定性高的時候,固定成本低和投資期限短,淨現值低,這是大多數人喜歡穩定的環境的原因。
在一個穩定的社會環境,大多數人的固定投資很高,尤其表現在教育方面,高等教育十分普遍,在一個不穩定的社會環境,象處於戰爭狀態的地區,大多數人的固定投資低,讀書的年數少。通常我們覺得,固定成本高和投資期限長是好事,但如果我們所在的地區, 突然變得很不穩定,可以計算出,固定投資越高的系統損失越大,這就是為什麼在社會大變動的年代,那些簡單的社會系統和底層的人反而能夠生存,在地質大變動的時代,那些處於統治地位的大物種,象恐龍,會滅絕,而簡單的物種反而能夠生存。 更具體的分析,請參考 Jing Chen and James Galbraith, A Biophysical Approach to Production Theory, Working paper
|



